The four bioindication methods (open water sandy cages, within-bed sandy cages, open water mesh cages, and within-bed mesh cages) were applied to investigate the environment condition suitability for FWPMs in the upper Vltava River Basin (Bohemian Forest, Czech Republic). This river represents one FWPM residual locality within central Europe19. Here, we present a specially selected set of results illustrating the most important aspects of the four methods. Further details are described in a comprehensive study by Černá et al.13.
The river environment was studied at two levels:
(I) A longitudinal river profile was represented by main stream localities (sites A – E) and tributaries of different pollution stages (sites R and V). The localities were tested both by sandy cages and by mesh cages installed in free-flowing water. In addition, a gravel hyporheic zone was tested by within-bed sandy cages in localities B, C, and D.
(II) A hyporheic environment was tested in the selected locality C. The suitability of different substrates (sand, gravel, stones) was tested by within-bed mesh cages.
The growth rate and survival rate of >1-year-old juveniles (see Supplementary File 1: S.6. Biological material) were tested. The experiment was carried out to its full extent in the summer of 2014 and was repeated to a smaller extent at some localities in the summer of 2015. Within level (I), 2 – 6 sandy cages with a minimum of 100 juveniles and 6 (2014) or 4 (2015) mesh cages with 6 juveniles were applied at each locality tested by the appropriate method. Within level (II), 7 mesh cages with 6 juveniles were installed in every tested environment. The exposure time was one month for the mesh cages and three months for the sandy cages.
The statistical analysis was conducted in R, version 3.1.020. Kruskal-Wallis, Kruskal-Nemenyi, and Wilcoxon-Mann-Whitney tests were used. For data with a normal distribution, linear or quadratic regression was carried out.
The localities can be clearly distinguished based on the growth rate in the open water mesh cages despite the high within-cage variability, even in different growth-favorable periods (Figure 3). In the more growth-favorable exposure in 2015 (growth rate 19.3 – 41.8%), a significant trend was discovered in the longitudinal profile where the growth rate increased downstream (Kruskal-Wallis test, p < 0.001). Importantly, the survival rate was equivalently high in both seasons (from 83%) (Figure 4A).
On the other hand, the open water sandy cages showed a different trend between the main stream localities in 2014: the growth rate increased downstream from locality A (52%) through the middle locality C (153%), and thereafter decreased again until locality E (46%) (a quadratic regression of absolute growth values: r2adj = 0.77, F2,13 = 25.66, d.f. = 16, p < 0.001). This trend was also confirmed in 2015 when the greatest growth rate was recorded at the middle locality C again. Also, the absolute growth rate values did not differ much between 2014 and 2015. On the other hand, the survival rate differed between the years, being much higher in 2015 (from 48% to 72%) than in 2014 (about 25%) (Figure 4B).
An effect of two different exposure methods is also clearly visible in the polluted tributary (locality V). The sandy cages exposed here during the three months showed 0% survival, while an 83% survival rate with some growth was recorded by open water mesh cages exposure here during the 30 days.
Results from the within-bed sandy cages illustrate different conditions in the hyporheic environment in comparison to open water in the relevant localities. The growth rate was always lower in the hyporheal sites than in open water, and the survival rate was much more variable (from almost 50% to 0%, Figure 4B).
A study of hyporheic microhabitats using the within-bed mesh cages showed a significant effect of the substrate composition on juvenile survival. The best conditions were recorded from the oxygen-saturated stony bottom (a survival rate close to 100%) while the worst (a < 40% survival rate) were indicated in poorly oxygenated sand where a very high variability in surviving was also detected. Hyporheic water oxygenation, which was repeatedly measured during the experiment, explains this trend (Figure 5).
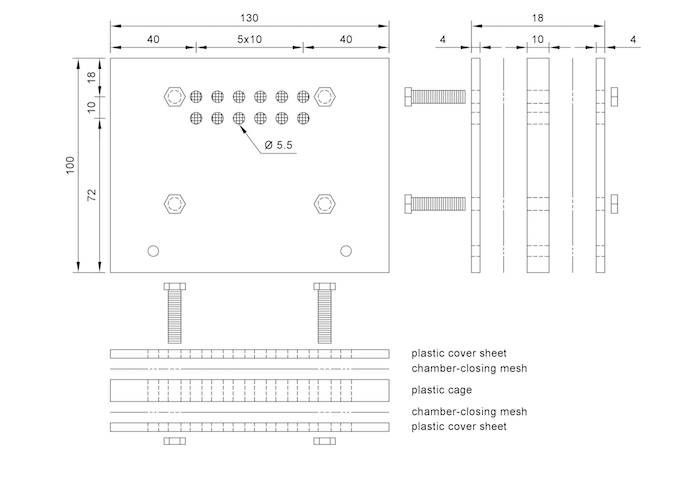
Figure 1. Bioindication mesh cage with individual chambers. See Supplementary File 1 for further details. Please click here to view a larger version of this figure.
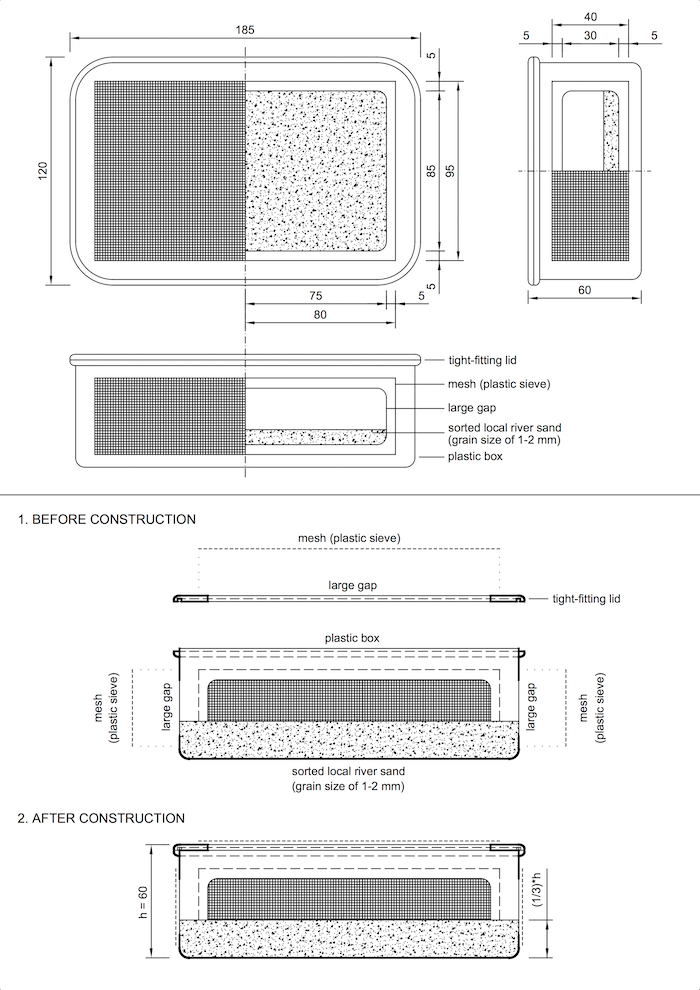
Figure 2. Bioindication sandy cage. See Supplementary File 1 for further details. Please click here to view a larger version of this figure.
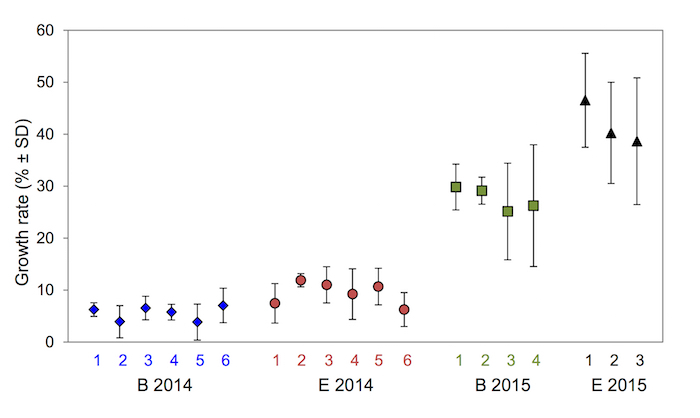
Figure 3. Individual variability in juvenile growth rate recorded by open water mesh cages in localities B and E during two seasons. The means and standard deviation are described for every mesh cage. The values are based on the measurement of 6 juveniles (or 4 – 5 juveniles if the mortality rate > 0%) in every mesh cage.
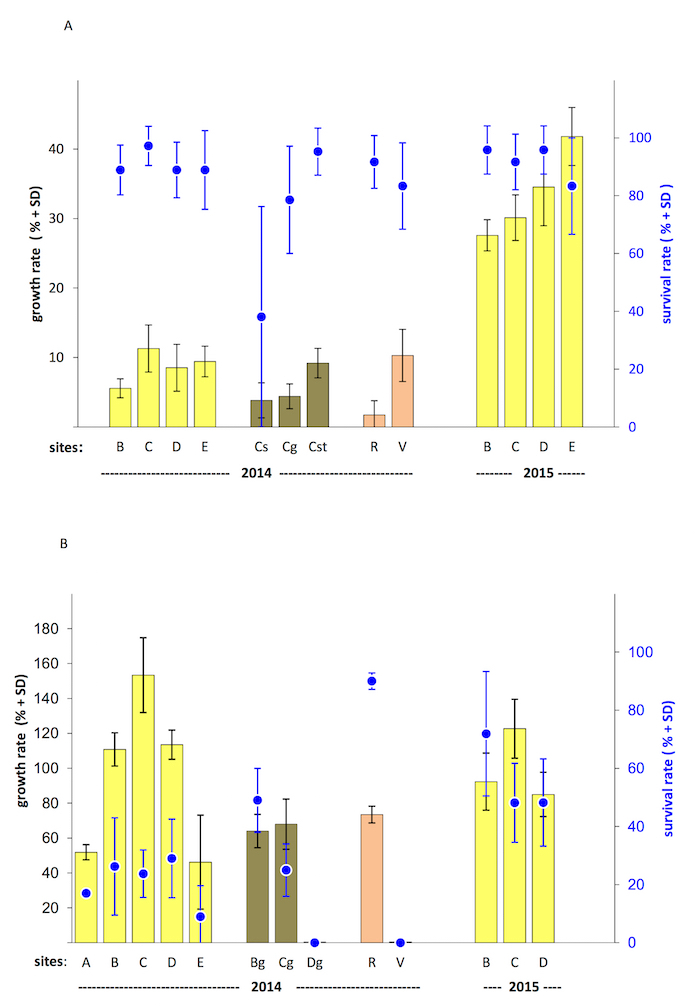
Figure 4. Example results from a field bioindication experiment with mesh and sandy cages. (A) This panel shows example results from a field bioindication experiment with mesh cages. A total of 6 localities (B, C, D, E, R, and V)within the Vltava River catchment were tested on 2 separate occasions (in 2014 and 2015). The exposure time was 30 days during the summer season. The Localities B – E represent (in order) a longitudinal profile of an approximately 20-km stretch of the river's main stream. Localities R and V represent profiles of 2 tributaries. Capitals mark the same locality both in panel (A) and (B). All localities were tested with open water mesh cages. In addition, locality C was also tested using within-bed mesh cages installed in 3 different types of river bed (Cs = sand, Cg = gravel, Cst = stones) in 2014. The cages were installed in 4 – 7 replications at every site. 6 freshwater pearl mussel juveniles of 1+ year old were used per mesh cage. The average growth rates are marked for the 3 largest individuals (3 MAX) from every tested mesh cage (columns, left axis) and the average survival rate per mesh cage (blue points, right axis). (B) This panel shows example results from a field bioindication experiment with sandy cages.A total of7 localities (A, B, C, D, E, R, and V)within the Vltava River catchment were tested over on 2 separate occasions (in 2014 and 2015). The exposure time was 3 months during the summer season. Sites A – E represent (in order) a longitudinal profile of an approximately 30-km long stretch of the river's main stream. Sites R and V represent profiles of 2 tributaries. Capitals mark the same locality both in this and in the previous panel. All localities were tested with open water sandy cages. In addition, localities B, C, and D were also tested using within-bed sandy cages installed in gavel river bed substrate (Bg, Cg, and Dg) in 2014. The cages were installed in 2 – 4 replications at every site. At least 100 freshwater pearl mussel juveniles were present in every sandy cage. The average growth rate for the 10 largest individuals (10 MAX) from every tested sandy cage (columns, left axis) and the average survival rate per sandy cage (blue points, right axis) are marked. Please click here to view a larger version of this figure.
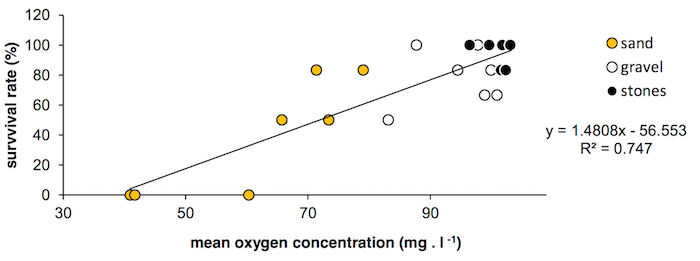
Figure 5. Oxygen saturation. This panel shows the relationship between minimal values of oxygen saturation over 30 days of mesh cages exposure and the surviving rate per cage in within-bed mesh cages exposed in different bed microhabitats in 2014. Please click here to view a larger version of this figure.
| 2014 | 2015 | |||
| locality | 3-month exposure of sandy cages | 1-month exposure of mesh cages | 3-month exposure of sandy cages | 1-month exposure of mesh cages |
| A | 13.9 | – | – | – |
| B | 14.4 | 13.4 | 13.9 | 17.5 |
| C | 15 | 13.8 | 14.4 | 18.3 |
| D | 15 | 13.8 | 14.3 | 18.3 |
| E | 15.5 | 14 | – | 18.7 |
| R | 13.5 | 12.8 | – | – |
| V | 14 | 13.2 | – | – |
Table 1. Average surface water temperature (°C) at the localities during exposure in 2014 and 2015.
