Necrotic cell death-induced by hyperactive ion channels
Using the procedures presented here, mec-4(u231) and deg-3u662) mutant embryos were either incubated for 25 min at 34 °C or kept at the standard temperature of 20 °C. Upon hatching, the number of neuronal cell corpses was determined at the L1 larval stage of both groups. Necrotic cell death is diminished in nematodes that hatched from heat shock preconditioned eggs (Figure 1A-1B).
Protein aggregate-induced neurodegeneration
Transgenic nematodes overexpressing (A) human α-synuclein and cytoplasmic GFP in dopaminergic neurons and (B) human polyglutamine protein (PolyQ) fused with YFP pan-neuronally, were exposed daily for 30 min at 34 °C. Heat shock preconditioning promotes neuroprotection against α-synuclein-induced cell death in 7-day-old adult hermaphrodites (Figure 2A) and decreases Q40::YFP protein aggregates in the head region of 4-day-old adults (Figure 2B and Figure 3).
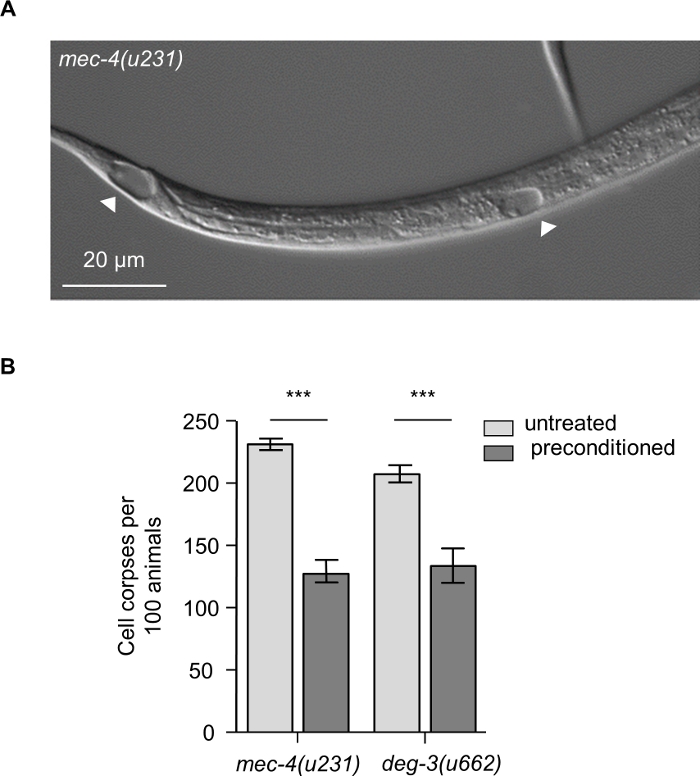
Figure 1: Hyperactive ion channel-induced necrosis. (A) Representative DIC microscopy image of mec-4(u231) L1 larva, with arrowheads indicating characteristic necrotic vacuoles. At early stage of neurodegeneration swollen nuclei are displayed within cells. Image was acquired by using a 40x objection lens. Scale bar, 20 μm. (B) Number of neuronal cell corpses at the L1 larval stage of development, per 100 animals carrying the neurotoxic mec-4(u231) or deg-3(u662) alleles. Necrotic cell death is suppressed in L1 larvae hatched from preconditioned eggs compared to untreated counterparts (n= 100 animals per genotype and assay; Data represent mean ±S.E.M., ***P< 0.001 for untreated versus preconditioned; t-test) Please click here to view a larger version of this figure.
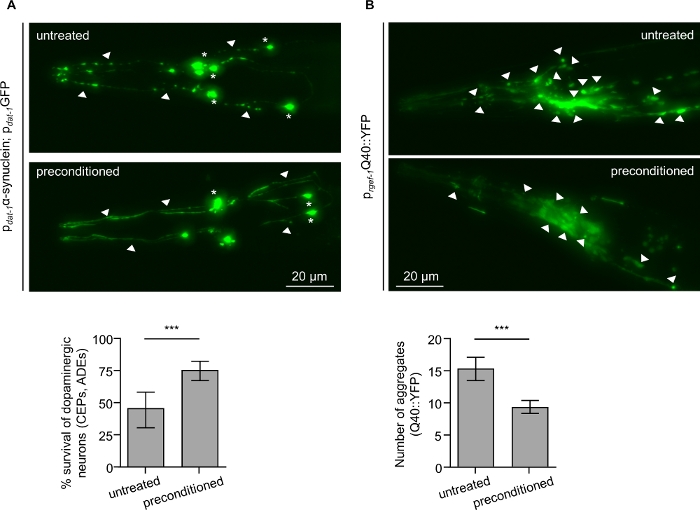
Figure 2: Daily preconditioning at 34 °C confers neuroprotection against α-synuclein and decreases polyQ aggregates in C. elegans. (Α) Survival of anterior dopaminergic neurons (CEPs and ADEs) in untreated and preconditioned nematodes co-expressing cytoplasmic GFP together with the human α-synuclein. Preconditioned nematodes display enhanced neuroprotection compared to untreated. Remnants of neuronal cell bodies (asterisks) and axonal beading (arrowheads) are seen in untreated nematodes (top panel). Both soma (asterisks) and neuronal processes (arrowheads) are preserved upon preconditioning. Images acquired by using a 20x objection lens and depict maximum intensity z-projection. Acquisition details: Bright, 0.15. Scale bar, 20 μm. Worms were scored for neuronal survival of the anterior dopaminergic neurons on the seventh day of adulthood. (B) Neuronal polyQ aggregates detected on the fourth day of adulthood in the head region of untreated and preconditioned transgenic nematodes. Preconditioned transgenic worms present less neuronal Q40::YFP aggregates compared to untreated. Representative images of the head region are shown with arrowheads indicating polyQ protein aggregates in neuronal cells. Images acquired by using a 20x objection lens and depict maximum intensity z-projection. Acquisition details: Bright, 0.0175. Scale bar, 20 μm. 30-35 animals were quantified per condition in each of three independent experiments. Data represent mean ±S.E.M., ***P < 0.05, unpaired t-test. Please click here to view a larger version of this figure.
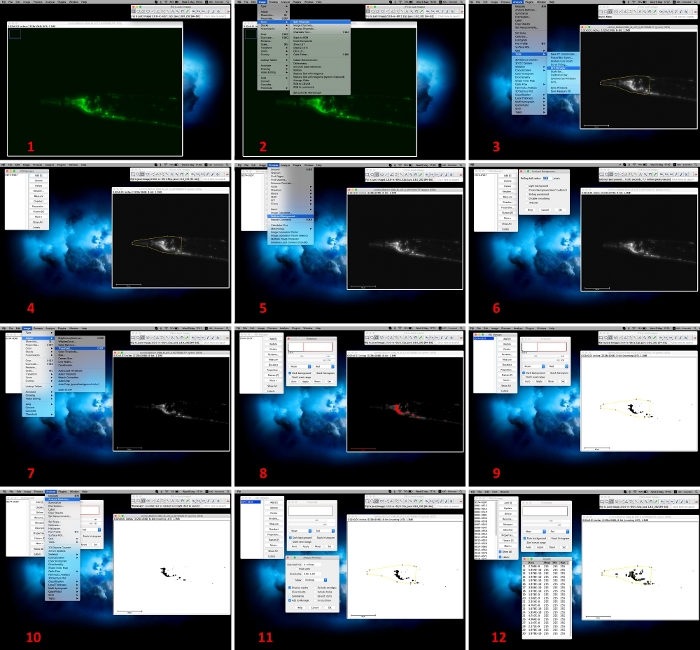
Figure 3: Image analysis by using Fiji software. 1. Open an acquired image with Fiji software; 2. Select “Split channel” command via the “Image” and “Color” drop down menu to convert the image; 3, 4. Keep “green channel” image and by using the “freehand selection” tool, enwrap the fluorescent region of interest (ROI; e.g., head). Add the respective ROI in “ROI Manager” via “Analyze” and “Tools” drop-down menu; 5, 6. Subtract the background to 50% by selecting “Process” and “Subtract Background”; 7, 8. Set up and apply threshold values via the menu command “Image”, “Adjust” and “Threshold”; 9. Select the respective ROI from the “ROI Manager”; 10-12. Analyze the number of protein aggregates by using the menu command “Analyze” and “Analyze Particles”. Please click here to view a larger version of this figure.
| Reagent | Recipe | |
| 2% agarose pads | 1. Weigh 0.5 g of agarose in a cylindrical glass beaker. | |
| 2. Add 25 mL of M9 buffer. | ||
| 3. Heat in a microwave until close to boiling. Take out, stir with a pipette tip and boil again. Repeat until the agarose is dissolved. | ||
| 4. Place an empty microscope slide on the bench. | ||
| 5. Put a drop (~ 50 µL) of fresh 2% agarose solution in the middle of the slide. | ||
| 6. Take a second microscope slide and place it on top of the agarose drop. Gently press down to flatten the drop. | ||
| 7. Let the agarose harden for 30 seconds and remove gently the top microscope slide. | ||
| 8. Immediately proceed with the sample preparation, since the agarose pads will start drying within approximately 5 minutes. | ||
| Tip: Leave the top microscope slide as a cover to preserve the humidity longer (~ 1 hour). Thus, several agarose pads can be prepared and used swiftly during the experiments. | ||
| M9 buffer | 1. Dissolve 3 g of KH2PO4, 6 g of Na2HPO4, 5 g of NaCl in 1 L of distilled water and autoclave. | |
| 2. Let cool and add 1 mL of 1 M MgSO4 (sterile). | ||
| 3. Store M9 buffer at 4 °C. | ||
| Nematode growth medium (NGM) agar plates | 1. Mix 3 g of NaCl, 2.5 g of bactopeptone, 0.2 g of streptomycin, 17 g of agar and add 900 mL of distilled water. Autoclave. | |
| 2. Let cool to 55-60 °C. | ||
| 3. Add 1 mL of cholesterol stock solution, 1 mL of 1 M CaCl2, 1 mL of 1 M MgSO4, 1 mL of nystatin stock solution, 25 mL of sterile 1 M phosphate buffer, pH 6.0, and distilled sterile water up to 1 L. | ||
| 4. Pipette 10 mL of medium per Petri dish and leave to solidify. | ||
| 5. Store the plates at 4 °C until used. | ||
Table 1: Recommended recipes for reagents used. All the reagents recipes used in the presented protocol are outlined here.
