After zebrafish larvae are properly immobilized and the posterior lateral line afferent ganglion and VR recording is achieved, activity in both afferent and motor neurons can be measured simultaneously. Recording channels are displayed using gap-free recording protocols (step 1.4) for continuous monitoring of afferent and VR activity. In real-time, decreases in spontaneous afferent spike rate can be observed concurrent with VR activity indicative of fictive swim bouts (Figure 1E). We found that best results and accurate spike detection were products of recordings that achieved a signal-to-noise ratio of at least 0.5. Custom written pre-processing scripts generate plots to assist in visualization of afferent and VR spike detection. Spontaneous afferent spikes are identified using a combination of spike parameters such as threshold, minimum duration (0.01 ms), and minimum inter-spike interval (ISI; 1 ms). Increasing negative pressure while establishing the recording often yields signal detection from multiple afferent units at once. Filtering by amplitude allows for distinguishing between signal dynamics of independent afferents. Isolating signals can be achieved by adjusting the lower-bound and upper-bound detection variables in the pre-processing script (Figure 2A). Aggressive suction to achieve multi-unit recordings can lead to unstable recordings, mechanical noise, degradation of afferent health, and ultimately a loss of signal. Therefore, it is important to slowly dial back suction to atmospheric pressure once the desired signal is achieved. Ventral root spike detection follows identical parameters to afferent spike detection but requires additional inputs to define distinct fictive swim bouts. Bursts within a motor command are defined by VR activity with a minimum of two spikes within 0.1 ms of each other and lasted a minimum of 5 ms. All swim bouts are then delineated by a minimum of three bursts with inter-burst intervals of <200 ms (Figure 2B).
Afferent activity is difficult to interpret when looking at a recording in its entirety. Pre-processing scripts will overlay sections of afferent activity centered on a well-defined period of interest, in this case, the onset of a swim bout (n = 33, Figure 2C) to assist in visualizing trends in signal dynamics. Instantaneous afferent activity is calculated using a moving average filter and a 100 ms sampling window. Mean spontaneous activity shows dramatic changes in response to the onset of motor activity (Figure 2C). To better dissect and analyze afferent activity, periods before and after the swim are set to match the time interval of the corresponding swim bout. In the pre-processing script and representative analyzed results these periods are termed "pre-swim" and "post-swim". Pre-swim, swim, and post-swim spike rates were calculated by taking the number of spikes within the respective period over its duration. The precision of estimates for each individual is partly a function of the number of swims, so we analyzed variable relationships using weighted regressions, with individual weights equal to the square root of the number of swims.
Differences in afferent spike rates across the various periods of interest (pre-swim, swim, and post-swim) were tested by a two-way analysis of variance (ANOVA). Tukey's post-hoc test detected significant differences in spike rates between swimming spike rates and spike rates of both pre-swim (8.94 ± 0.2 Hz, relative decrease 57%) and post-swim (5.34 ± 0.2 Hz, relative decrease 40%) periods. The spike rate did not immediately return to the baseline given we also found that post-swim spike rate was lower than the pre-swim spike rate (Tukey post-hoc tests across groups, p < 0.001; Figure 3A). Linear models were used to detect relationships between relative spike rate and fictive swim parameters. Relative spike rate was calculated by taking the swim spike rate over the pre-swim spike rate. Fictive swim parameters included swim duration, swim frequency (i.e., number of bursts within a swim bout over the duration of the swim bout), and duty cycle (i.e., sum of the swim burst durations over the swim bout total duration). In our hands, the mean and variance of relative spike rate was correlated, so it was necessary for the data to be log transformed for analysis. Afferent spike rate was negatively correlated with swim duration meaning that the lateral line experiences greater inhibition during swims of longer duration (r2 = 0.186, F2,26 = 2.971, p = 0.045; Figure 3B). There was no correlation detected between relative spike rate and neither swim frequency nor duty cycle ((r2 = 0.099, F2,26 = 1.431, p = 0.231, and r2 = 0.047, F2,26 = 0.645, p = 0.932, respectively; Figure 3C,D). All analyses of variable relationships were weighted by the number of swims per individual and all the variables were then averaged by each individual (n = 29).
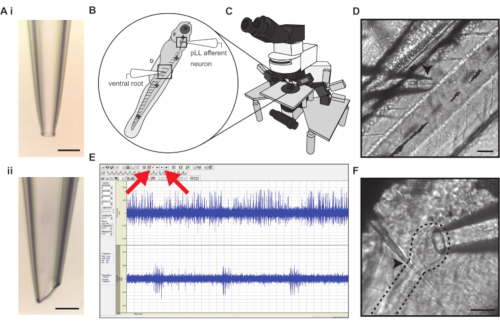
Figure 1: Simultaneous electrophysiological recording of posterior lateral line afferent neuron and ventral motor root activity. (A). Example of a loose-patch afferent (i) and ventral motor root (ii) recording electrodes. Scale bars represent 50 µm. (B) Larval zebrafish are paralyzed and pinned in four locations (cross symbols) to a Sylgard dish for recording stability. Bold crosses represent insertion points for pins. (C) The electrophysiology rig is mounted on a vibration-isolation table and consists of an upright fixed stage microscope on a motorized controller capable of 40x magnification. Dual current clamp and voltage clamp head stages are mounted on micromanipulators. (D) The myomeres of the body musculature are separated by myosepta that serve as recording landmarks for motor neuron arborizations. The ventral motor root electrode approaches the ventral body (left) and is centered and lowered on top of a myoseptum (arrowhead). Scale bar represents 50 µm. (E) Screen capture of electrophysiological recording in real-time allowing visualization of the spontaneous afferent activity (channel 1) and bursting ventral root activity indicative of fictive swim bout (channel 2). The Record and Play buttons are denoted with red arrows. (F) The posterior lateral line afferent ganglion (dashed line) lies just under the skin and can be identified by a tight cluster of afferent soma. The ganglion can be located by following the lateral line nerve past the cleithrum bone (arrowhead) to where it connects to the ganglion. Scale bar represents 30 µm. Please click here to view a larger version of this figure.
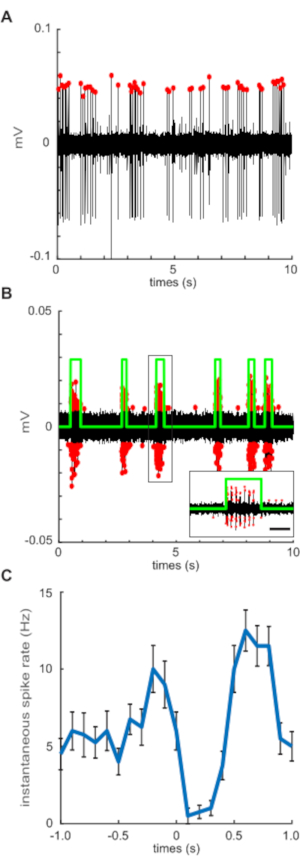
Figure 2: Pre-processing figure outputs visualize accurate spike detection. (A) Extracellular, loose-patch recording of posterior lateral line afferent neurons. Discrete spikes (labeled with red dots) are detected with a minimum inter-spike interval of 1 ms. Baseline noise and activity from other units are filtered out by thresholding to only include spike amplitudes within 50% of the maximum. (B) Ventral motor root recording (VR) of fictive swim bouts reveal voluntary motor commands throughout the duration of the recording. VR spikes (red dots) are detected using a similar threshold filter and then binned into a single swim bout (green) by detecting a burst of activity within 200 ms of one another (see insert; scale bar represents 200 ms). Spikes detected outside the defined swim bout do not occur within the inter-spike interval of stereotyped burst activity and are therefore excluded. (C) Mean spontaneous afferent spike rate centered on the onset of each swim bout (time = 0 s) illustrating spike rate before, during, and after swimming. Error bars represent ± SEM. Please click here to view a larger version of this figure.
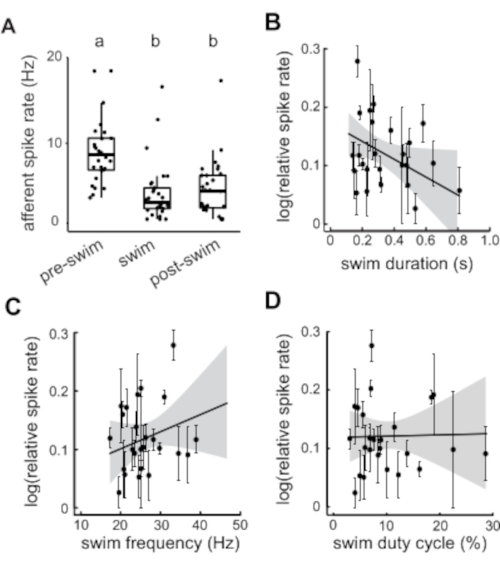
Figure 3: Quantification of afferent activity before, during, and after fictive swimming. (A) Afferent spike rate is significantly reduced during swimming and this effect persists even afterwards. Statistically similar groupings are denoted by a and b. (B) Longer swim duration is correlated to decreased afferent spike rate. (C–D) Swim frequency and swim duty cycle show no correlation to afferent spike rate. All values represent mean ± SEM. Outlying individuals with low statistical weight were omitted. Please click here to view a larger version of this figure.
