1. Protocol Text
This protocol highlights the adaption of NMR methodology to M. tuberculosis (Class III agent). Therefore, Biosafety Level 3 (BSL3) practices need to be followed when conducting M. tuberculosis research in an annually certified laboratory. Exposure to laboratory-generated aerosols is the most important hazard encountered by personnel working with these microorganisms. The following procedures are conducted at our institution and variations may exist based on the Institutional Biosafety Committee’s recommendations. Common personnel protective equipment will consist of a Tyvek suit, bouffant cap, booties, N95 respirator, eye protection, sleeves, and a double pair of nitrile gloves. Work involving M. tuberculosis cultures and/or manipulations with open M. tuberculosis containers is performed in the type A2 or B2 biological safety cabinet. Plastic-covered absorbent paper is placed on the working surface. All materials/supplies to be disposed or removed from the facility must be placed in two biohazard bags and decontaminated by autoclaving. Work surfaces and equipment utilized within the cabinet (FastPrep-24 lysis homogenizer, spectrophotometer, bucket with ice, etc.) must be disinfected after each work session with 1% Amphyl (tuberculocidal, bactericidal, fungicidal, and virucidal agent). M. tuberculosis cultures must be placed under double containment for transportation to larger equipment located outside of the biological safety cabinet such as freezers, incubators, centrifuges, and refrigerators. Centrifugation is conducted with enclosed safety cups and O-ring screw top tubes. For further analysis outside the BSL3 laboratory, cell-free extracts are filtered through a 0.2 μm filter or microorganisms heat killed at 95 °C for 15 min.2 Samples are plated to verify the absence of colony-forming units prior to removal from containment.
- Transfer 110 ml of Middlebrook 7H9 complete media containing polysorbate 80 (Tween 80 to prevent clumping) broth (MADC-TW) or a suitable medium to a 250 ml Erlenmeyer flask. Triplicate cultures are routinely grown per condition when using 13C-metabolites, and ten identical cultures are grown for 1D 1H NMR profiling. For two conditions (e.g., with and without addition of drug), double volume of culture per replicate can be grown and split into two identical cultures. All reagent and solution recipes are provided at the end of the protocol.
- Inoculate broth with 0.150 ml of M. tuberculosis 50% glycerol stock (allow to thaw on ice). See Note 1 below.
- Let the culture grow at 37 °C shaking at 100 rpm for approximately 6 days (OD600 0.6-0.8). See Note 2 below.
- If no antibiotics, alternative additions or other treatments are needed, the cultures are ready to be harvested. If treatments are used, continue directly to Step 5. Remove a 0.5 ml sample from each flask, transfer to a microcentrifuge tube and place at 4 °C for culture titration and quality control: contamination tests, phenotypic analysis, PCR tests. See Note 3 below. At this point, proceed to Step 6.
- Remove flasks from shaker and perform the desired treatment (e.g., add drug or metabolite). Place flasks back in the shaker and incubate for an additional period (e.g., 6-18 hr). At the end of this time, take another 1.0 ml aliquot and determine OD600. Remove a 0.5 ml sample from each flask, transfer to a microcentrifuge tube and place at 4 °C for culture titration and quality control.
- Place cultures on ice for 5 min. After this step, leave cells on ice throughout the entire remaining protocol. Harvest cultures by centrifugation at 2,000 x g and 4 °C for 15 min in 50 ml tubes using a benchtop centrifuge. Each culture will require four tubes with 25 ml each (a total of 100 ml is required to obtain adequate signal-to-noise for 2D 1H-13C NMR experiments, where only 50 ml per culture is required for 1D 1H NMR experiments).
- Wash each cell pellet two times with ice-cold ddH2O (approximately 15 ml the first time and 10 ml the second time) by the centrifugation parameters described above. For the second wash, combine the 10 ml aliquots into the same tube before spinning. Resuspend the cell pellet in a final volume of 1.0 ml ddH2O (need to adjust for cell pellet volume). If a single cell suspension free of clumping is desired at this step, cells can be briefly sonicated and/or passed through a 27-gauge needle three times. Alternatively, the cell pellet can be frozen at -80 °C and stored until further processing. In the latter case, frozen pellets are thawed on ice prior to resuspension. Resuspend the cell pellets in a final volume of 1.0 ml ddH2O as stated above.
- Transfer the 1.0 ml cell suspension to a 2.0 ml screw cap tube containing Lysing Matrix B (0.1 mm silica spheres). Place the FastPrep-24 lysis homogenizer inside the biosafety cabinet. Put the samples into the sample holder, securing the retention spoke plate on top of the tube. Process the samples for 60 sec in the homogenizer at a speed setting of 6 meters/sec.
- Spin the samples in a microcentrifuge at 15,000 x g and 4 °C for 10 min to pellet cell debris and intact cells.
- Remove supernatant and pass sample through a syringe filter (0.2 μm) into a sterile tube. Plate 0.1 ml of sample (or a representative fraction of the sample, as for example 10%) on MADC agar to verify that there are no viable cells. Investigators may also conduct more than one filtration step and/or verify the presence of viable cells in the extracts using live-dead staining procedures so as to prevent or identify any potential biosafety concerns. Freeze samples in an ethanol-dry ice bath to store at -80 °C until they are ready to be lyophilized and processed in an NMR facility.
- After 2 months, check plates to verify the absence of colony forming units. If no CFU’s are found, samples may be taken out of the BSL3 laboratory. Further analysis is carried out in a standard NMR facility, which normally serves multi-users and operates under no specific containment requirements.
- Lyophilize the samples to dryness, then resuspend in 0.7 ml of NMR buffer and transfer to a microcentrifuge tube. Centrifuge for 3 min at 13,000 x g. Remove 0.6 ml and transfer to a 5 mm NMR tube. Alternatively, lyophilized samples could be mailed to an external facility as non-biohazardous regular samples.
- NMR data is collected immediately. Despite all precautions, active enzymes may still be present and the sample is typically not stable for long periods of time. Changes in the NMR spectrum are noticeable when the samples are left out at room temperature or at 4 °C for more than 1 week. Also, NMR data collection is alternated between untreated and drug treated cultures, where samples are randomly selected from each category. This prevents unnecessary bias because a particular category had a longer delay before the NMR spectra were collected. A bias in the data would occur if the NMR spectra for all the untreated samples were collected first, and then followed by the drug treated samples. If the samples are unable to be immediately analyzed by NMR, the samples should be stored in 1 ml Eppendorf tubes at -80 °C.
- After inserting the samples into a BACS-120 sample changer, instrument calibration, which involves a few routine steps such as lock, shim, tune, and optimization of the 90 ° pulse length, is required to maximize the quality of the results. A single sample is used to calibrate the 90 ° pulse length and tune the instrument for the remaining samples. Locking, shimming, and NMR data collection for every sample is automated using ICONNMR and gradshim.
- A 1D 1H NMR spectrum is collected using the Bruker zgesgp pulse sequence with water suppression using excitation sculpting. A total of 32k data points with a sweep width of 5482.5Hz, 128 scans, and 16 dummy scans are used. A 2D 1H-13C HSQC spectrum is collected using the Bruker hsqcetgp pulse sequence. A total of 2048 data points with a sweep width of 5000.0 Hz is collected along the direct 1H dimension, and 64 data points with a sweep width of 18864.9 Hz along the indirect 13C dimension. The spectrum is collected with 128 scans and 16 dummy scans to obtain good signal to noise.
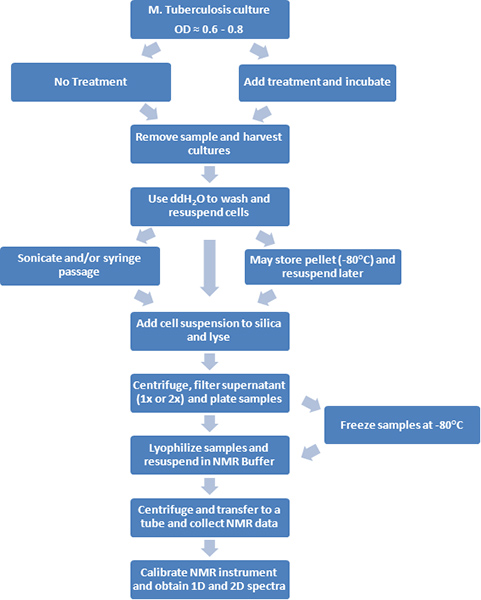
Figure 1. A flow diagram of the experimental procedures is depicted.
NMR buffer
Stock solution of 50 mM potassium phosphate buffer with 50 μM TMSP:
- 2.17 g K2HPO4 (potassium phosphate dibasic)
- 1.70 g KH2PO4 (potassium phosphate monobasic)
- 7.86 mg Sodium-3-Trimethylsilylpropionate (TMSP-2,2,3,3-D4 (D,98%) )
- Dissolve in 500 ml of “100%” D2O
- The final solution should be at a pH 7.2 (uncorrected)
MADC-TW or MOADC-TW (1 L)
- 4.7 g Middlebrook 7H9 broth base
- 900 ml ddH2O
- 2 ml glycerol
- Mix the components and autoclave for 25 min. Cool liquid to touch and add the following solutions:
- 100 ml ADC when preparing MADC or 100 ml OADC when preparing MOADC
- 2.5 ml 20% Tween 80 (alternatively, 1 ml of non-metabolizable 20% Tyloxapol)
- 10 ml 1% cycloheximide
ADC (1 L)
- 20 g D(+)-Glucose
- 50 g BSA Fraction V
- 8.5 g NaCl
- 800 ml ddH2O
- Dissolve together, adjust volume to 1 L and sterilize with 0.2 μm filter. Store at 4 °C.
Alternatively, to prepare OADC (1L), add all components listed above plus 50 ml of 1% oleic acid (recipe below makes a 250 ml batch). Dissolve together, adjust volume to 1 L and sterilize with 0.2 μm filter. Bottle needs to be wrapped in aluminum foil and store at 4 °C.
- 2.5 g Oleic Acid
- 250 ml 0.2M NaOH
- Thaw oleic acid (solidifies upon refrigeration) by heating at 55 °C for 10 min and add to NaOH solution and heat with stirring for 60 min. Store in sterile glass bottles that are wrapped in aluminum foil. Seal bottles with parafilm and store at 4 °C.
If preferred, you can purchase commercial BD BBL Middlebrook ADC or OADC with catalase Enrichment.
1% Cycloheximide (100ml)
- 1 g cycloheximide
- 100 ml ddH2O
Dissolve and sterilize with 0.2 μm filter. Store at 4 °C. Caution: cycloheximide is toxic so handle with extreme care.
20% (v/v) Tween 80 (100ml)
- 20 ml Tween 80
- 80 ml ddH2O
Alternatively, weigh 20 g of Tween 80 (approximately 18.9 ml; density 1.06 g/ml), to prepare 20% w/v solution. Heat solution to 55 °C for 30 min to dissolve, and mix completely. Sterilize liquid with a 0.2 μm filter. Store the final solution at room temperature.
20% (v/v) Tyloxapol (100ml)
- 20 ml Tyloxapol
- 80 ml ddH2O
Alternatively, weigh 20 g of Tyloxapol (approximately 18.2 ml; density 1.1 g/ml), to prepare 20% w/v solution. Heat solution to 55 °C for 30 min to dissolve, and mix completely. Sterilize liquid with a 0.2 μm filter. Store the final solution at room temperature.
Note 1: Glycerol stocks of M. tuberculosis are prepared using the following protocol.
Stocking of M. tuberculosis
- Grow M. tuberculosis in 50 ml MADC to saturation (OD600 = 1.5 to 2.0) at 37 °C under shaking conditions (100 rpm). Depending on the strain, this will take 7-14 days.
- Spin samples at 2,000 x g at 4 °C for 15 min to pellet culture. Remove supernatant.
- Resuspend in 6 ml of sterile 50% glycerol.
- Aliquot 1.5 ml into 4 Corning cryogenic vials and label appropriately.
- Immediately flash freeze vials in ethanol/dry ice bath and store at -80 °C.
Note 2: Inoculating strain H37Rv (-80 °C stock up to 1.5 years old) into 70 ml of MADC media yields an OD600 of about 0.6 after 5 days of growth at 37 °C under shaking conditions (100 rpm) in an Innova 40 Shaker.
Note 3: To test culture contamination, investigators could inoculate a culture aliquot onto standard rich medium and verify absence of growth after overnight incubation. Routinely, cultures are plated on MADC agar to observe colony morphology and examine by phase-contrast microscopy. If desired, cultures can be verified by PCR using IS6110 primers as described.3
2. Representative Results
A sample that is well prepared will yield an NMR spectrum similar to the one depicted in Figure 2. This spectrum is a representation of the M. tuberculosis wild type metabolic pool. The identified metabolites are directly or indirectly associated with the D-alanine pathway. Also, the magnitude of peak intensities is proportional to metabolite concentrations present in the cell extract. Therefore changes in peak intensities between untreated cultures, drug treatment, and mutant strains can indicate perturbations in metabolites and metabolic pathways. The M. tuberculosis 1D 1H NMR spectra were collected on a Bruker Avance 500 MHz spectrometer equipped with a triple-resonance, Z-axis gradient cryoprobe. The spectrum contains ca. 400 peaks, from which it was possible to identify and quantify 40-50 metabolites, including amino acids, nucleotide precursors, and glycolytic and citric acid intermediates. A BACS-120 sample changer with Bruker Icon software was used to automate the NMR data collection. 1D 1H NMR spectra were collected using excitation sculpting4 to efficiently remove the solvent and maintain a flat baseline, eliminating any need for baseline collection that may induce artifacts in subsequent principal component analysis (PCA) or orthogonal partial least squares discriminant analysis (OPLS-DA). A 1D 1H NMR spectrum is collected at 25 °C with a spectrum width of 5482.5 Hz and 32K data points. A total of 16 dummy scans and 128 scans were used to obtain the spectrum. The ACD Labs 1D NMR processor software was used to semi-automatically process all the 1D 1H NMR spectra. The spectra were Fourier transformed, phased, and referenced to the TMSP peak (0.0 ppm). The NMRpipe software package was used to individually process the 2D 1H-13C NMR spectra and analyzed with NMRDraw. The Bruker FID data file was first converted to a file format recognizable by NMRpipe and then the spectrum was Fourier transformed, phase corrected, and zero filled. The observed NMR peaks in the 1D 1H NMR spectrum and 2D 1H-13C NMR spectrum are assigned to specific metabolites using 1H and 13C chemical shift tolerances of 0.05 and 0.50 ppm, respectively, and the Madison Metabolomics Consortium Database (MMCD),5the BioMagResBank,6 and the Human Metabolome Database.7 Specifically, the 1D and 2D NMR spectra are manually peak-picked, where the peak list of NMR chemical shifts is then uploaded to the Human Metabolome Database. The metabolites identified by the Human Metabolome Database are assigned to the NMR spectrum based on both maximizing the number of matching NMR resonances and belonging to a metabolic network. Each compound or metabolite typically has multiple C-H pairs and correspondingly multiple NMR resonances. Thus, the more of these NMR resonances that are observed in the experimental NMR spectrum, the more likely the metabolite is present. Similarly, identifying multiple metabolites associate with the same pathway increases the likelihood of correct assignments. The presence of metabolites and metabolic pathways are verified with the Kyoto Encyclopedia of Genes and Genomes (KEGG)8 and the MetaCyc databases.9 Mycobacterium smegmatis is a useful model system for M. tuberculosis and other mycobacterial pathogens. As described elsewhere, samples of M. smegmatis could also be disrupted by sonication.10
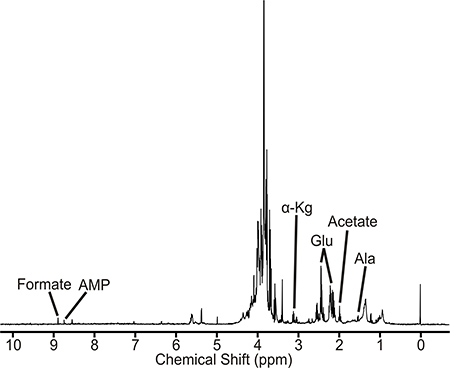
Figure 2. 1D 1H NMR spectrum of Mycobacterium tuberculosis cell extract. NMR peaks associated with representative metabolites are labeled. Abbreviations are the following: AMP, adenosine mono phosphate; α-Kg, α-ketoglutarate; Glu, Glutamate; and Ala, alanine.
