To ensure a high islet yield during isolation, surgical techniques outlined in the protocol should be followed closely. Although the techniques presented here will be tailored to each laboratory, there are a few critical steps that will lead to a successful isolation. In order to make the common bile duct easily accessible, it is recommended that the organs be displaced to the right side of the mouse (Figure 1). Moreover, this will allow the pancreas to inflate with a smaller amount of resistance since there will be less weight restricting expansion. Another critical step to maximize islet yield is the ligation of the common bile duct close to the Sphincter of Oddi (Figures 2A-C). Ligating further away from the Sphincter of Oddi may result in a partial inflation by reducing the amount of collagenase solution entering the major pancreatic duct. Also, a taut knot will prevent collagenase solution from entering the intestine.
A proper incision into the common bile duct should be located far enough away from the bifurcation from the liver but close enough for maximal pancreatic inflation (Figure 3). An incision made too close to the bifurcation may lead to the flow of collagenase solution into the liver. When collagenase solution enters the liver, it will lose its dark-red color and begin to turn whitish. If this occurs, remove the needle and try another cannulation further from the liver. Additionally, the common bile duct incision should only be part way through the duct (about 50%). Completely shearing the duct will make it difficult to seal the duct around the needle tip. A proper incision will result in complete closure of the duct around the needle, producing enough pressure to both fill the common bile and pancreatic ducts (Figure 4A). It is also important to ensure the needle has entered the common bile duct and not the surrounding sheath. Once the needle has been inserted properly, 5 ml of the collagenase solution should fill the pancreas, creating a marbleized tissue.
The pancreas removal process should be performed delicately to promote maximal islet yield (Figure 5). Piercing the pancreas may result in a deflation and loss of collagenase solution, reducing the islet yield. Cutting the pancreas close to the connective tissue and other organs will prevent deflation. Also, harvesting other tissue, such as connective tissue, along with the pancreas is not an issue, as it will be removed in subsequent steps. A properly removed pancreas will remain inflated after excision prior to further collagenase digestion (Figure 6).
Distinct layers in the Ficoll gradient will lead to a clean islet preparation and create an easier environment for picking islets (Figure 7). The gradient ensures the separation of islets from cell debris and connective tissue that remains after digestion and mesh screen filtering. Furthermore, this step is crucial for selecting clean, healthy islets for downstream applications. Specifically, healthy islets will be spherical in shape and have a golden brown to dark brown center (Figure 8). Note: Islets from diabetic mice will be much paler in color, corresponding with decreased insulin content, and can be better distinguished from acinar tissue by their shape, glossiness, and their visible network of capillaries. Any islets connected to acinar tissue should not be used and should be discarded. Moreover, larger islets that develop a dark, necrotic center after an overnight incubation should be discarded as they do not function properly.
As described in the introduction, cAMP production is an important component of β-cell function; in particular, with regards to incretin action. One benefit to the protocol described in this method is that one can simultaneously obtain cAMP production and glucose-stimulated insulin secretion data. Typically, the impact of an agent on cAMP production directly correlates with its impact on insulin secretion17. For simple experiments, this rule holds true fairly well (see Figure 9 for an example). In the present protocol, we use IBMX (a phosphodiesterase inhibitor) in our treatments to prevent the catabolism of cAMP, giving the total production of cAMP.
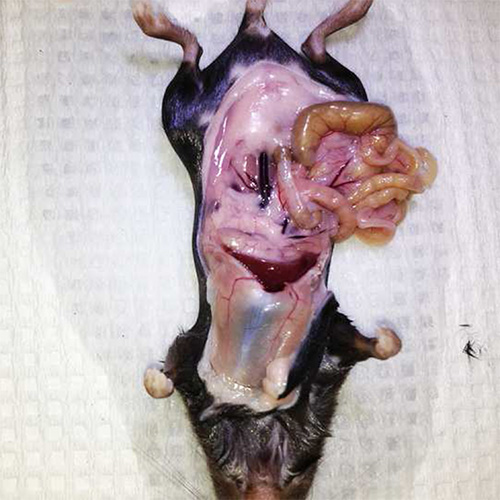
Figure 1. Displacing internal organs. Positioning the internal organs to the right side of the mouse creates an easier work environment and allows the pancreas room to expand during inflation.
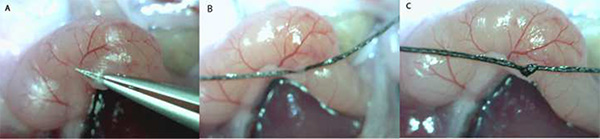
Figure 2. Tying off the Sphincter of Oddi (A-C). Depicted here is the entrance of the common bile duct to the small intestine at the Sphincter of Oddi. Tying off the common bile duct at the Sphincter of Oddi prevents collagenase solution from entering the intestines.
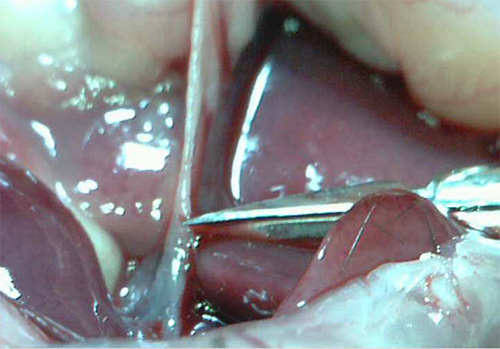
Figure 3. Common bile duct incision. It is best to make a cut in the common bile duct slightly distal to the bifurcation into the liver to prevent flow of the collagenase solution into the liver.
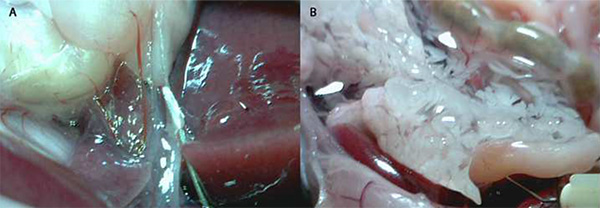
Figure 4. Cannulation of the common bile duct by a blunted 30 G needle and pancreas inflation. A) The 30 G needle is inserted into the common bile duct, creating a seal around the needle tip. It is important to check and to note the needle has been inserted into the common bile duct and not the surrounding sheath. A properly inserted needle will have an opaque appearance. B) A proper pancreas inflation will fill with about 3-5 ml of collagenase solution and have a marbled appearance.
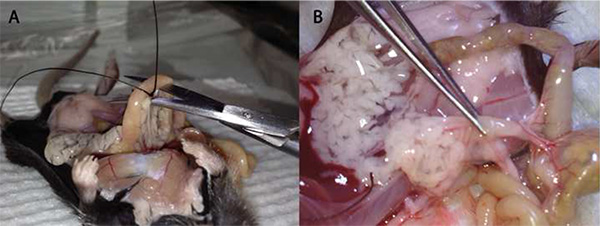
Figure 5. The removal of the pancreas. A) The initial incision made with a curved pair of scissors occurs near the Sphincter of Oddi. The initial removal begins in the direction of the stomach being careful not to puncture the pancreas. B) The final step is removing the pancreas from the connective tissue on the small intestine and the last few connections to the peritoneal cavity.
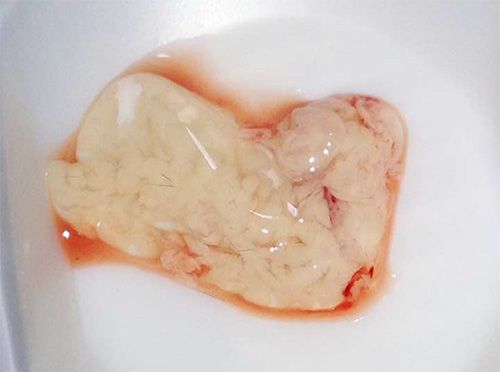
Figure 6. The inflated pancreas. A successful pancreas removal will yield a pancreas perfused with the collagenase solution. Poor excision will leave a smaller, deflated pancreas.

Figure 7. Four layers of the Ficoll gradient. The four distinct layers represent a different Ficoll density from 25% at the bottom to 11% at the top. The distinct layering is imperative to remove debris and exocrine disuse from the islets.
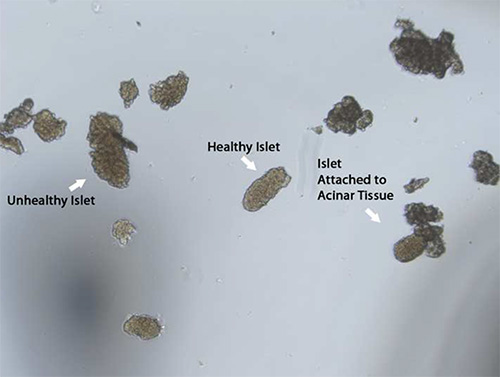
Figure 8. Islet selection. Healthy islets tend to have a golden brown to dark brown color with a round spherical shape and are not connected to acinar tissue. After an overnight incubation (16-20 hr) a necrotic center develops in larger islets, which should be excluded from experiments.
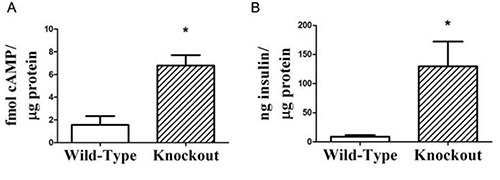
Figure 9. Representative high-quality cAMP production results, demonstrating that secreted insulin can be measured from the cAMP stimulation medium. A) Isolated islets from wild-type or gene knockout mice were stimulated with 11.1 mM glucose and intracellular cAMP production was measured and normalized to the total cellular protein. B) Secreted insulin was measured using a standard insulin ELISA, and also normalized to total cellular protein. In many cases, the change in cAMP production correlates directly with the augmentation in glucose stimulated insulin secretion. n=3 for each group; *, p < 0.05. This figure was adapted from research published in Kimple et al10.
