Results are shown in Figure 2. A single lateral group (LG) terminal cell shows extensive subcellular branching (visualized by GFP; A) and a gas-filled subcellular lumen running through each of the branches (visualized by brightfield microscopy; B). These images were collected from a mosaic L3 larva, generated using the MARCM system described in sections 1 & 2, and heat fixed and imaged, as described in sections 3 & 4. Panels C & D show a NeuronJ generated trace, as described in section 5, of the branches and the lumen respectively of the images shown in A & B. Panels E & F show the location of the fat body (FB) branch in a larva prepared for imaging as described in sections 3 & 4. Note that in this example, the animal is not a mosaic, and GFP is expressed throughout the entire tracheal system. Panels G and H show an example of a GFP-labeled terminal cell that was heat fixed for too long a time. GFP is diffuse (G), branches have broken down and portions of lumens are no longer gas filled, thus appearing as breaks in the brightfield image.
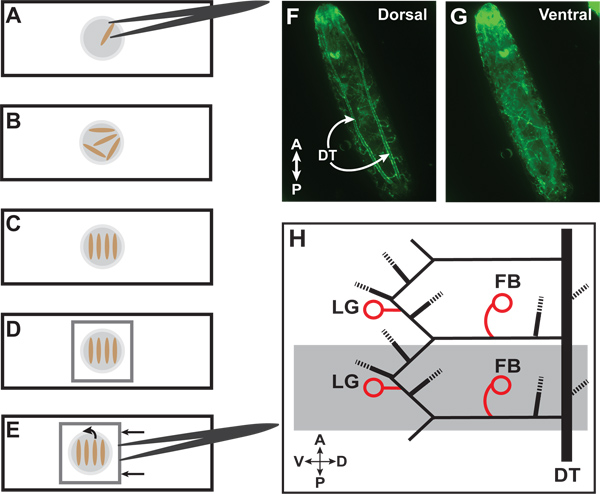
Figure 1. Larvae preparation and identification of terminal cells. (A) Place larva in a drop of 100% glycerol on a glass slide. (B) Multiple larvae can be placed for heat fixation. (C) After heat fixation, organize larvae parallel to each other and perpendicular to the long axis of the slide. (D) Place cover glass over larvae. (E) To reorient larvae, carefully push cover glass with forceps to roll the animals. (F) Wild-type third instar larva with GFP expressed throughout the tracheal system. The paired dorsal trunks (DT) are visible on the dorsal side. (G) The same larva after the rolling technique with the ventral side now facing upwards. (H) Diagram of lateral view of two third-instar tracheal hemisegments (one hemisegment is highlighted in grey). Circles indicate lateral group (LG) and fat body (FB) terminal cells which we use for quantitation. Dashed lines represent other branches of the tracheal system which we do not routinely quantitate. For a full description of the tracheal branches in a larval segment, refer to Ref 1.
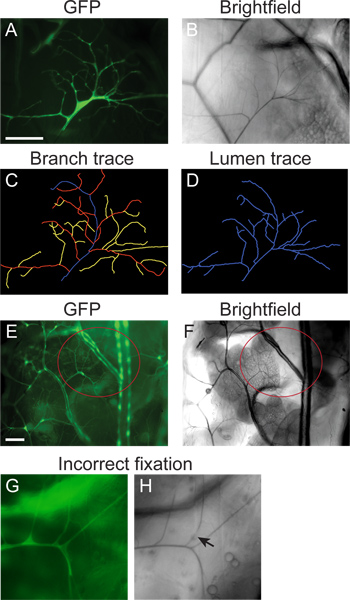
Figure 2. Representative images. (A-D) Mosaic L3 larvae were generated using the MARCM technique (section 1) and fixed and imaged using the protocol in sections 2-4. The branching pattern of a single LG terminal cell was visualized by mosaic expression of GFP (A); the gas-filled lumen visualized with brightfield microscopy (B). (C) Tracing of the branching pattern of the cell in A, generated using NeuronJ (section 5). (D) Tracing of the gas-filled lumen of the cell in B, generated using NeuronJ (protocol section 5). (E, F) A fat body (FB) terminal cell (highlighted by the red circle) visualized by GFP expression throughout the tracheal system (E) and brightfield microscopy (F) in a larva prepared by the protocol in sections 2-4. (G, H) Example of fixation artifacts obtained when the sample is heated for too long a period. GFP is diffuse and small branches have degraded (G). Areas of lumen also no longer appear air-filled (arrow in H). Scale bar: 100 μm.
