In order to obtain reliable, reproducible results using a neutrophil binding assay, it is essential that the health and the confluency of the microvascular endothelial cells are optimal on the day of the assay, as illustrated with HMVEC-Lung in Figure 1. In addition, it is imperative that low passage number microvascular ECs are used (i.e. less than 9 passages), and accordingly, we recommend performing all experiments within two weeks of thawing. The health of the neutrophils is also of utmost importance, especially since calcein AM will not be metabolized to the fluorescent calcein molecule if the cells are comprised in some manner. We use the trypan blue exclusion method, and do not proceed with the assay if a significant proportion of the cells are dead (i.e. stained).
There are several ways to isolate neutrophils, including density gradient and antigen-based column separation methods, and any of these should suffice for this procedure. We chose to use the Polymorphprep ready-made density gradient solution from Axis-Shield. As can be seen in Figure 2, the PBMCs are present in the top layer, while the neutrophils are within the lower layer after centrifugation. The PBMC layer is aspirated in order to avoid contamination of the granulocytes upon their removal. It is important to note that step 2.9, in which water is added to lyse any remaining red blood cells, is optional. However, while it is ideal to remove all contaminating erythrocytes, it is critical that the neutrophils do not sit in pure water for more than the 30 sec after which considerable death can occur.
Finally, the neutrophils are incubated with calcein AM for 30 min, washed and resuspended in pre-warmed (37 °C) RPMI-1640 media (without phenol red). It is important to use media without phenol red, which can interfere with the fluorescence reading. The number of neutrophils added to the wells is rather arbitrary, provided that the fluorescence readings pre- and post-wash are within the linear range of the spectrophotometer, and the differences between treatments that either promote or prevent neutrophil adherence can be reproducibly ascertained (see below). We find that the fluorescence OD520nm is within the linear range when 10,000 to 1,000,000 labeled neutrophils are analyzed (Figure 3A). Interestingly, we also find that the percent adherence increases as more neutrophils are added to HMVEC-Lung monolayers treated with TNFa (100 ng/ml) for 3 hr (Figure 3B). We chose to incubate the HMVEC-Lung and calcein-labeled neutrophils together for 20 min, since we find that the percentage of adhesion does not significantly increase after this time point (data not shown). Of note, in this assay we used standard, clear polystyrene 48-well plates and found that the level of fluorescence cross-over readings between the wells did not amount to more than 0.5% of the total fluorescence input (data not shown). Nonetheless, we recommend using 48-well plates purposely made for fluorescence spectrophotometer readings for more sensitive experiments, if available, or at the very least, designing your experiment to minimize cross-over readings (see Step 1.2).
To demonstrate that the number of neutrophils added to the wells is rather arbitrary so long as the number added is within the linear range of the spectrophotometer, we performed a neutrophil adhesion assay with TNFα-treated HMVEC-Lung in the absence or presence of the p38-MAPK inhibitor BIRB7906, using varying input amounts of neutrophils (i.e. 40,000/well, 200,000/well or 1,000,000/well) 25. p38-MAPK has previously been shown to be necessary for E-selectin expression in TNFα-treated human ECs, and its inhibition should therefore reduce neutrophil binding to TNFα-activated HMVEC-Lung 26. In fact, this is what we observe (Figure 3C). Although we find that the percent adherence increases as more neutrophils are added, and the percent adherence may differ between experiments, due to various factors such as inter-donor variations, the quality of the neutrophils and the density of HMVEC at confluence, the relative adherence between conditions remains relatively constant in each independent experiment regardless of the number of neutrophils added (Figure 3C and data not shown). These data also exemplify why it is better to display the adhesion data as relative to a positive control for inter-experimental consistency.
In order to fully illustrate how this assay works, we performed a neutrophil adhesion assay with TNFα-treated HMVEC-Lung pre-incubated with either control antibody or an E-selectin blocking antibody. E-selectin is expressed on the EC surface after treatment with TNFa and captures neutrophils from the vasculature via electrostatic interactions with glycomolecules present on the neutrophil surface 1,27 . As shown numerically in Figure 4A, graphically in Figure 4B and visually in Figure 4C, the TNFα-treated HMVEC-Lung pre-incubated with the E-selectin antibody bound ~75% less neutrophils than the TNFα-treated HMVEC-Lung pre-incubated with control IgG. This suggests E-selectin plays an important role in tethering neutrophils to HMVEC-Lung in our static adhesion assay.
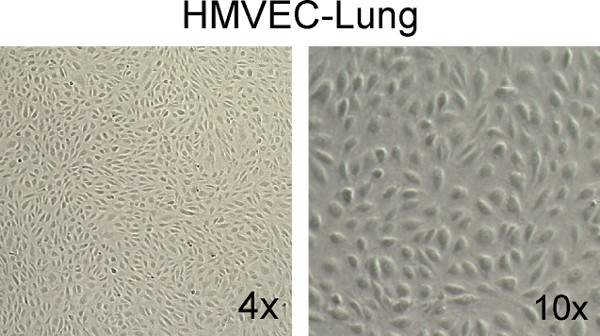
Figure 1. Example of healthy, confluent HMVEC-Lung monolayers. Note the "cobblestone" appearance and total confluency of the monolayers. Images were taken at 4X and 10X magnification on a Fisher Scientific Micromaster inverted digital microscope.
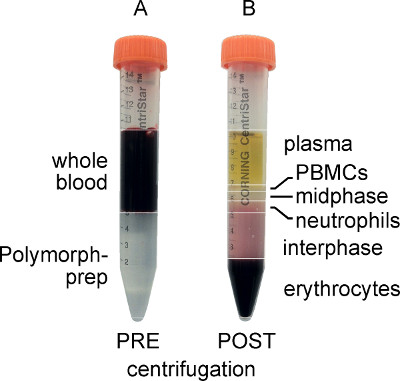
Figure 2. Whole blood and Polymorphprep layered prior to centrifugation (A) and separated after centrifugation (B). Note that after centrifugation, the PBMCs form a distinct upper band, while the polymorphonuclear granulocytes (neutrophils) form a distinct lower band.
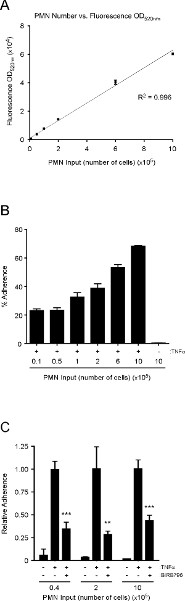
Figure 3. The relationship between neutrophil number and fluorescence OD520nm is linear; the percent adhesion of neutrophils to TNFα-treated HMVEC-Lung increases as more neutrophils are added; p38-MAPK promotes neutrophil adherence to TNFα-activated HMVEC-Lung monolayers. (A) Graph depicting the linear relationship between fluorescence OD520nm and neutrophil numbers over two orders of magnitude. An R2 value of 0.996 indicates a linear relationship between neutrophil number and fluorescence OD520nm when 10,000 to 1,000,000 neutrophils are analyzed. (B) Graphical representation of the percent adherence when varying numbers of calcein-labeled neutrophils were added to untreated or TNFα-treated (100 ng/ml; 3 hr) HMVEC-Lung monolayers in 48-well plates. (C) HMVEC-Lung monolayers were pre-incubated with the p38-MAPK inhibitor BIRB796 (10 mM) (Axon Medchem) or DMSO (control) for 1 hr prior to TNFa (100 ng/ml) treatment for 3 hr while in the continuous presence of BIRB796 (10 mM). The relative adherence was calculated as in step 4.8. Data are expressed as the mean ± SD. GraphPad Prism unpaired t-test was used for statistical analysis (n = 3) (TNFα-treated vs. TNFα-treated plus BIRB796). p-value ** < 0.01; *** < 0.001. Click here to view larger figure.
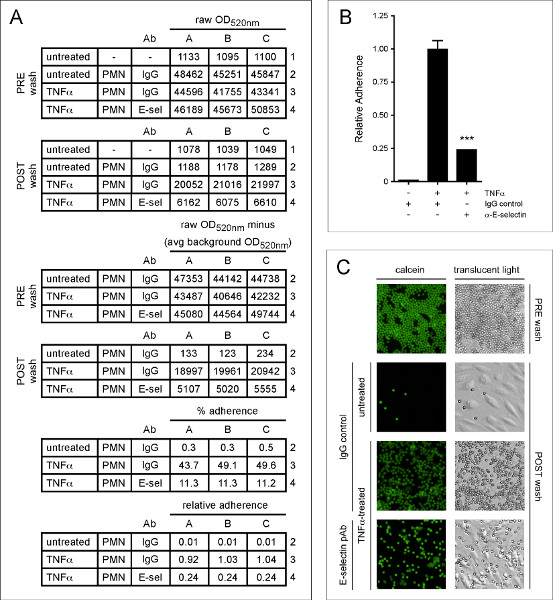
Figure 4. E-selectin promotes the adherence of neutrophils to TNFa-treated HMVEC-Lung. HMVEC-Lung were treated with TNFα (100 ng/ml) for 3 hr. After washing the HMVEC-Lung with RPMI 1640 containing 3% BSA (step 4.1), either sheep IgG (50 μg/ml) (5-001-A; R&D Systems) or E-selectin polyclonal antibody (pAb) (50 mg/ml) (AF724; R&D Systems) were added to the appropriate wells for 20 min. The HMVEC-Lung monolayers were then washed once more with RPMI-1640 before 6 x 105 neutrophils were added per well for an additional 20 min. (A) The calculations to determine the percent and relative adherence are shown. Calcein light emission is measured at OD520 nm. Background OD520 nm average is derived from HMVEC-Lung in the absence of TNFα treatment and neutrophils. (B) Graphical representation of relative percent adherence of calcein-labeled neutrophils to untreated or TNFα-treated HMVEC-Lung monolayers after pre-incubation with either control IgG or E-selectin blocking pAb. Data are expressed as the mean ± SD. GraphPad Prism unpaired t-test was used for statistical analysis (n = 3). Calculated p-value was < 0.001. (C) Images of calcein-labeled neutrophils bound to untreated and TNFα-treated HMVEC-Lung monolayers pre-incubated with either control IgG or E-selectin pAb. An example well containing 6 x 105 neutrophils prior to washing with PBS is shown (PRE wash). Translucent light microscopy images are shown in the right-hand column, while fluorescence images of the same field of reference, using a fluorescein filter set, are shown in the left-hand column. Images were taken at 10X magnification on an Olympus IX51 inverted microscope equipped with a Retiga 2000R camera (Q Imaging). PRE wash = total neutrophil fluorescence OD520nm (Step 4.4); POST wash = adherent neutrophil fluorescence OD520nm (Step 4.7); PMN – polymorphonuclear granulocytes; avg – average; IgG – sheep IgG control; E-sel/α-E-selectin – E-selectin blocking pAb. Click here to view larger figure.
