A comparison of Protein A column purified Nurr1-specific antibodies with Protein A purified anti-Nurr1 antibodies followed by passage through Nur77 LBD and Nor1 LBD columns is shown in Figure 1. As can be seen, Protein A-purified Nurr1 antibody exhibited a strong binding to Nurr1 LBD. However, it also showed significant binding to Nur77 LBD and to Nor1 LBD. When Protein A-purified Nurr1 antibodies were further purified against Nur77 LBD and Nor1 LBD, the final affinity purified Nurr1 antibodies exhibited specific binding to Nurr1 LBD with undetectable binding to Nur77 LBD or Nor1 LBD, demonstrating that its cross-reactivity to Nur77 and Nor1 was completely removed.
This specificity was further demonstrated by Western blot analysis of extracts from CHO cell transfected with expression vectors carrying full length Nurr1, Nor1, or Nur77 fused to Myc tagging protein (Figure 2). As expected, anti-myc antibodies revealed that each full-length protein was expressed at its expected molecular weight. In agreement with the ELISA results, Protein A-purified Nurr1 specific antibody robustly detected full-length Nurr1 but also detected Nor1 and Nur77 although less efficiently. In contrast, the fully purified Nurr1 antibody did not exhibit any detectable cross-reactivity to Nor1 and Nur77 by ELISA or Western blot analyses.
Finally, the fully purified Nurr1 antibody was used for immunohistochemical analysis of mDA neurons. It is well documented that Nurr1, but not its close homologs Nor1 and Nur77, is prominently expressed in rodent and human mDA neurons at both mRNA and protein levels14,15. The use of the fully purified Nurr1 antibody confirmed that Nurr1 is almost exclusively expressed in the nucleus of mDA neurons in the substantia nigra area (Figure 3).
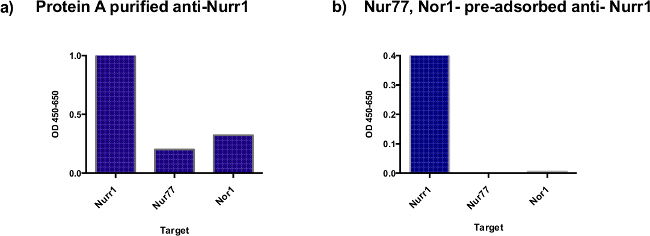
Figure 1: ELISA comparison of cross-reactivities of anti-Nurr1 antibodies before and after purification by passage through Nur77 and Nor1 LBDs coupled columns. Protein A purified anti-Nurr1 antibodies (0.156 μg/ml) (a) or protein A purified followed by chromatography onto Nor1 and Nur77 LBD-coupled columns anti-Nurr1 antibodies (0.156 μg/ml) (b) were added to wells of 96 well plate coated with 100 μl of 2.5 μg/ml of either Nurr1 LBD, Nor1 LBD or Nur77 LBD. The ELISA was carried out as described in the method section.
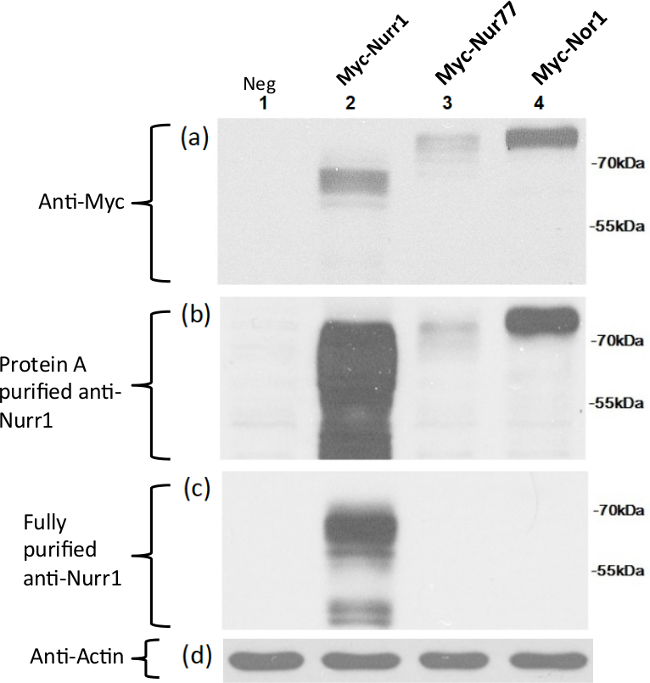
Figure 2: Specific detection of full-length Nurr1 expressed in CHO cells using the fully purified Nurr1-specific antibody. Full-length Nurr1, Nor1, and Nur77 proteins were expressed as myc tagged fusion proteins in CHO cells and detected by Myc specific antibodies (a), Protein A-purified Nurr1 antibody (b), and fully purified Nurr1 antibody (c). Anti-actin antibodies used for protein loading control (d). Each Myc-tagged full length DNA (molecular weight of 65, 66, and 69 kDa for Nurr1, Nur77, Nor1, respectively) was transiently transfected into CHO cells and the same amount of cell extracts were loaded in each lane. Lane 1: negative control consisting of CHO cells extracts transfected with an empty vector; lane 2: Myc-Nurr1; lane 3: Myc-Nur77; lane 4: Myc-Nor1.
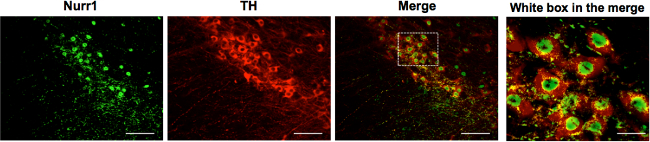
Figure 3: The fully purified Nurr1-specific antibody specifically detects tyrosine-hydroxylase-positive dopamine neurons. Midbrain dopamine neurons in the substantia nigra are positive for Nurr1, as examined by immunohistochemistry using the fully purified Nurr1-specific antibodies. Notably, Nurr1 was localized in the nucleus of mDA neurons. TH (tyrosine hydroxylase). Scale bar = 100 μm. Scale bar in white box merge is 10 μm. Please click here to view a larger version of this figure.
