As seen previously during the protocol description, the experiment needs 5 day to complete (Figure 1: experiment timeline). One operator is solicited during the entire run of the experiment and can handle the processes up to a maximum of 10 mice. If more animals are required, two persons are needed particularly for surgical sample collection. Indeed all samples must be collected in under 2 hr to avoid an increased passive alveolar-capillary leakage of FITC-labeled albumin in the last mice.
The first step is the preparation of C. albicans inoculum and intra-nasal instillation to obtain the airway colonization by C. albicans. A 4 days-persistence model is obtained by intranasal instillation of 5 x 105 CFU of C. albicans per mouse (Figure 2B). During these 4 days, mice gain weight (Figure 2A) and instillation of 5×105 CFU does not induce lung injury (Figure 2C). Although C. albicans may persist up to 4 days in this model, load decreases after 48 hr. Therefore, P. aeruginosa-induced acute lung infection is perfomed at 48 hr of C. albicans persistence.
P. aeruginosa strain PAO1 is a largely characterized laboratory strain comprising the major virulence factor, the type three-secretion system (T3SS), as in 75% of clinical lung isolates 15. For theses reasons, PAO1 is a relevant strain in animal models of acute lung infection. Lung injury is assessed through alveolo-capillar permeability measured by protein leak from the vascular compartment into the airway expressed as the lung injury index. Lung injury increases with the inoculum (Figure 3A). Here we report kinetics of the acute lung injury component of our model induced by PAO1 strain (5×106 CFU/mouse) (Figure 3B-3F) alone without prior C. albicans-mediated priming. Depending on strain and time course of the model, the choice of initial P. aeruginosa inoculum is discussed in the next section and is suggested in Table 1.
Five mice per group were used. Lung injury index (Figure 3B), bacterial burden in the lung (Figure 3C), bacterial burden in the spleen, reflecting bacterial dissemination (Figure 3D), BAL cellularity (Figure 3E) and differential cell count (Figure 3F) were determined every 12 hr. Lung injury was maximal between 24 hr and 36 hr after infection (Figure 3B). Bacterial burden showed a 1-log CFU/ml decrease every 24 hr (Figure 3C). Cumulative bacterial dissemination assessed by spleen homogenate cultures increased each day (Figure 3D). Finally, While BAL cellularity in uninfected mice is mainly composed (90%) of alveolar macrophages, in BAL from infected mice, neutrophils were widely recruited and differential cell count showed 90% neutrophils and 10% macrophages and lymphocytes (Figure 3E, 3F).
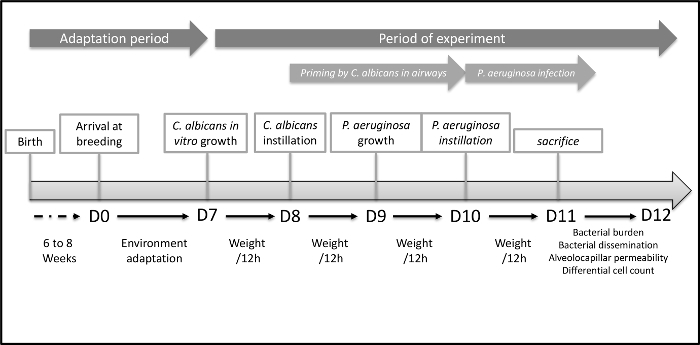
Figure 1. Timeline of the Acute Lung Injury Model to Explore Host-mediated Interaction between C. albicans and P. aeruginosa.
Graphic representation of the entire procedure. The first step is environmental adaptation of mice the housing facility. The Second step is C. albicans mediated airway colonization. Finally, the third step is the acute lung infection mediated by P. aeruginosa. Please click here to view a larger version of this figure.
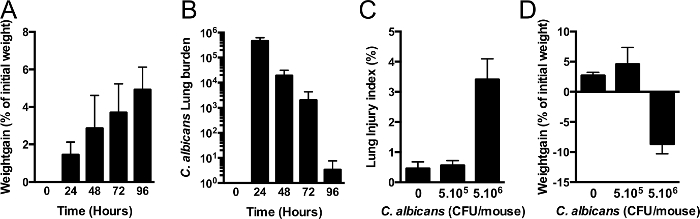
Figure 2. C. albicans airway Colonization.
(A,B) Mice are intranasally instilled with 105 CFU C. albicans (strain SC5314). Mice gain weight during C. albicans-mediated airway colonization (A). Colonization of the airway can be prolonged to 3-4 days with only one initial instillation. In a previous study, priming of innate immunity takes place between 24 and 48 hr. (n = 5 per group), error bars represent means ± SD. (C,D) mice are intranasally instilled 5 x 105 or 5 x 106 CFU of C. albicans. Lung injury index (C) assessed by alveolar capillary barrier permeability at 24 hr. Weight gain (D) expressed as percent of initial weight (n=5 per group). Error bars represent means ± SD. Please click here to view a larger version of this figure.
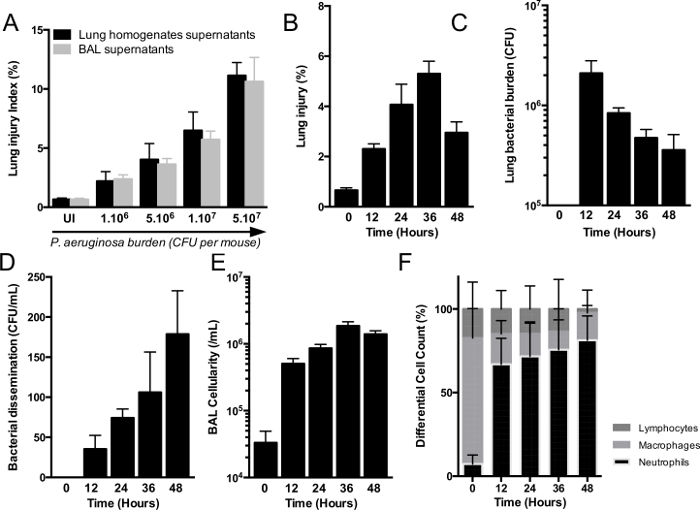
Figure 3. Model of Acute Lung Injury Induced by P aeruginosa.
(A) C57Bl/6J mice are intra-nasally infected with increasing loads of P. aeruginosa (from 1 x 106 to 5 x 107 CFU per mouse) (n=5 per group), error bars represent means ± SD. Mice are euthanized at 24 hr. Lung injury index is assessed by alveolar-capillary barrier permeability that increases proportionally with bacterial burden. Comparison of lung injury index obtained using old method with lung homogenates supernatants (black bars) and new combined-method using bronchoalveolar lavage supernatants (grey bars) (B-F) mice are intra-nasally infected with 5 x106 CFU per mouse. Mice are euthanized every 12 to 48 hr to acute injury model kinetics. Lung injury (B), lung bacterial burden (C), spleen bacterial burden (D), bronchoalveolar lavage (BAL) cellularity (E) and BAL differential cell count (F) are also assessed. (n=5) per group, error bars represent means ± SD. Please click here to view a larger version of this figure.
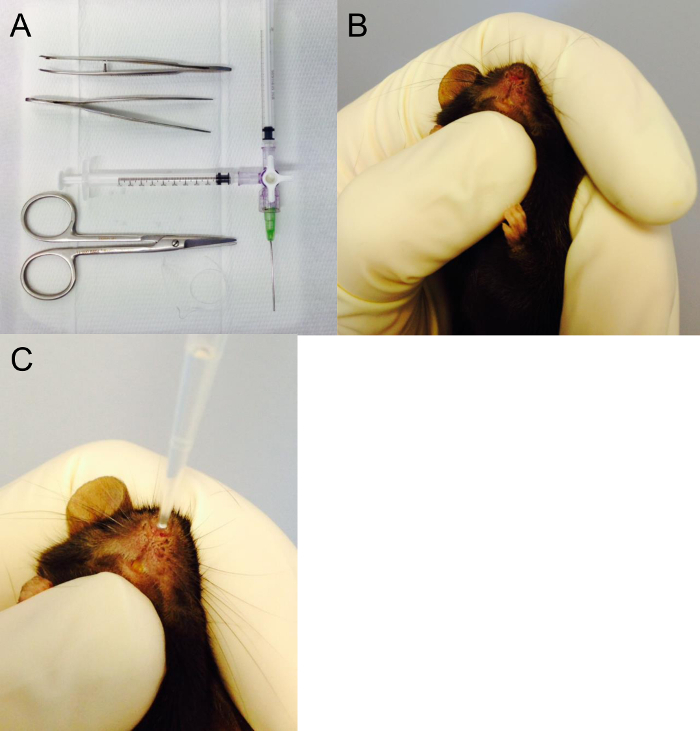
Figure 4. Surgical Equipment and Intranasal Instillation.
(A) Surgical equipment required to perform acute injury model and bronchoalveolar lavage. Here Tracheal cannula (20 G) and the two 1 ml syringes are connected to a Luer-lock 3-way valve. One syringe to inject water into the lungs, one to draw the bronchoalveolar fluid back out from the lungs. (B,C) Position of the mouse in the hand to perform the intra-nasal instillation. In this photo, the thumb under the jaw ensures a closed mouth during instillation. Please click here to view a larger version of this figure.
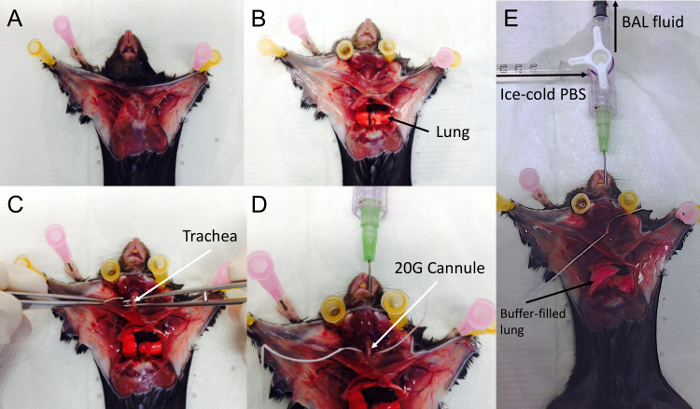
Figure 5. Surgery and Broncho-alveolar Lavage.
The chest is widely opened (A), and rib cage is opened laterally to avoid injury to the heart (B). Following blood collection, the cervical area is dissected to expose the trachea (C). Dental floss is used as a suture and is passed behind trachea (C,D). The trachea is then cannulated with the 20-G cannula combined (D) mounted on the syringe and 3-way valve. The trachea should be tightly secured around the cannula by tying a surgical knot using the suture in place behind the trachea. Finally 500 µl of PBS are gently instilled in the lungs and then the BAL is gently drawn out. (E) Fluid-instilled lungs. Please click here to view a larger version of this figure.
| Minimal Instilled Burden | Maximal Instilled Burden | |
| T3SS- | 5 x 107 | 1 x 108 |
| T3SS+ | 5 x 106 | 1 x 107 |
| T3SS+ exoU+ | 5 x 104 | 1 x 105 |
Table 1. P. aeruginosa Inocula Used in Acute Lung Infection Models.
Suggested optimal intranasal concentrations of inocula to induce acute lung injury according to strains.
