To determine the specificity of HDAC1,2-selective inhibitors, Hdac1,2FL/FL and Hdac3FL/FL fibrosarcoma cells were used. Adenovirus-containing Cre recombinase (Ad-Cre) was used to delete Hdac1,2 and Hdac3 in these cells. Following Ad-Cre infection of Hdac1,2FL/FL cells, a robust increase in H4K5ac was observed. Treatment of Hdac1,2 knockout cells with 233 or 898 did not result in any further increase in H4K5ac confirming that 233 and 898 inhibit Hdac1,2 and not Hdac3 in these cells. In contrast, addition of 233 or 898 to Hdac3 knockout cells resulted in a significant increase in H4K5ac compared to the increase seen in Hdac3 knockout cells due to an additive effect on H4K5ac levels as a result of inhibition of Hdac1,2 and 3. Hence, 233 and 898 are Hdac1, 2-selective inhibitors. These results are shown in Figure 1.
To validate the BrdU-ChIP-Slot technique, BrdU-pulse chase analysis was performed to look at the kinetics of PCNA loading on to the nascent DNA in HeLa cells. Our results showed that PCNA association with nascent DNA occurs rapidly within 15 min and disappears after a 30 min chase in agreement with previously published results13. These results are shown in Figure 2.
BrdU-H4K16ac ChIP-Slot-Western technique was used to determine the amount of H4K16ac associated with nascent DNA in the absence of HDAC1,2 function. A robust enrichment in H4K16ac associated with nascent DNA was observed when compared to the rabbit IgG control (Figures 3A and 3B). The increase in nascent DNA-associated H4K16ac following inhibition of HDAC1,2 activities or knockdown of Hdac1,2 is shown in Figure 3C, 3D, and 3E.
The level of SMARCA5 chromatin remodeler on nascent DNA was determined using BrdU-SMARCA5 ChIP-Slot-Western technique. Our results showed that SMARCA5 associates with nascent DNA in mammalian cells. Furthermore, HDAC1,2 inhibition or knockdown of Hdac1,2 did not change the amount of nascent DNA-associated SMARCA5 chromatin remodeler as shown in Figure 4.
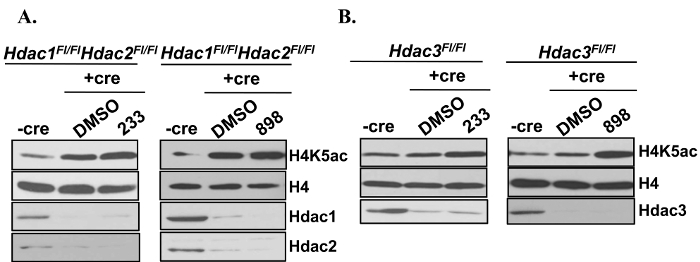
Figure 1. Confirmation of the Specificity of HDAC1,2-selective Inhibitors. Western analysis of whole cell lysates prepared from Hdac1FL/FL Hdac2FL/FL or Hdac3FL/FL fibrosarcoma cells following Ad-Cre infection and treatment with 898 or 233. Cells were treated with 3 µM 898 or 233 for 24 hr following a 48 hr Ad-Cre infection. This figure is derived from our previous published work6. Please click here to view a larger version of this figure.
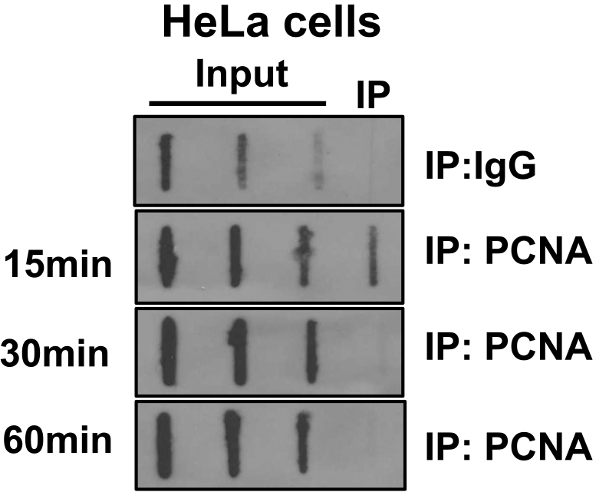
Figure 2. Dynamics of PCNA Association with the Nascent DNA in HeLa Cells. HeLa cells were labeled with bromodeoxyuridine (BrdU) for 30 min. Cells were then washed to remove unincorporated BrdU and cultured in media without BrdU for indicated periods of time (chase). Chromatin immunoprecipitation (ChIP) was performed with anti-proliferating cell nuclear antigen (PCNA) antibody. BrdU labeled DNA present in input DNA and those associated with PCNA were assessed in slot blot analysis using an anti-BrdU antibody as shown in Figure 2. This figure is derived from our previous published work6. Please click here to view a larger version of this figure.
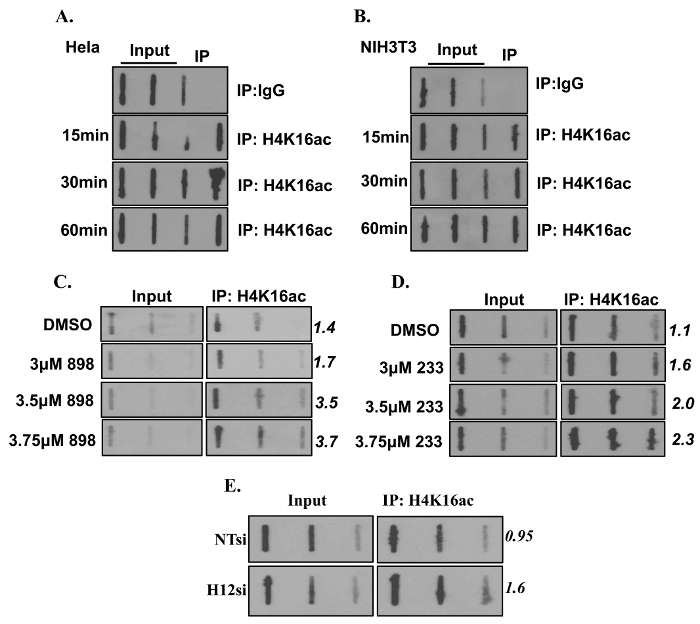
Figure 3. Loss of Histone Deacetylase 1 and 2 Increases H4K16ac on Nascent DNA. (A-B) Bromodeoxyuridine (BrdU) pulse chase was performed in HeLa and NIH3T3 cells to determine association of H4K16ac with nascent DNA at the indicated time points. (C-D) NIH3T3 cells were treated with either DMSO or a HDAC1,2-selective inhibitor (898 or 233) for 24 hr. (E) NIH3T3 cells were either transfected with non-targeting (NT) or Hdac1,2 (H12) siRNA. Cells from (C – E) were labeled with BrdU and used for ChIP with anti-H4K16ac followed by Slot blotting. The membrane was probed with anti-BrdU antibody. Numbers indicate to Image J quantitation of the average BrdU signal of high and medium volume of ChIP DNA spotted. This figure is derived from our previous published work6. Please click here to view a larger version of this figure.
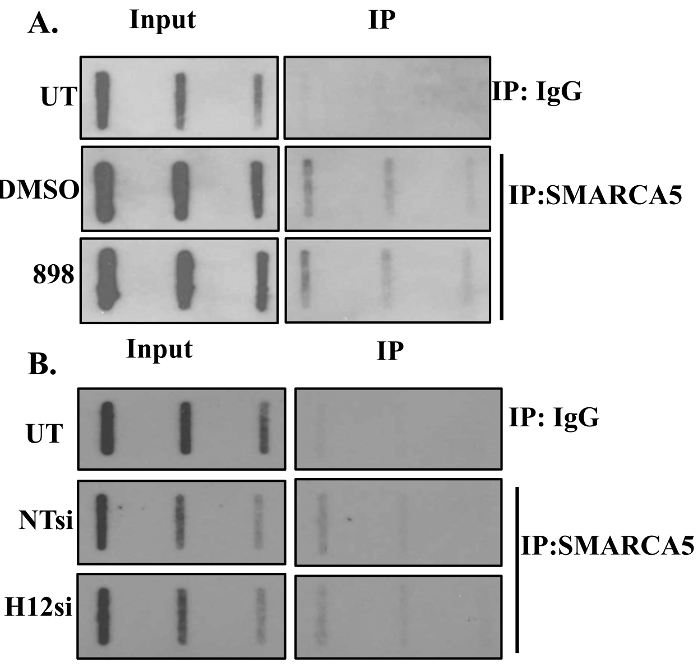
Figure 4. SMARCA5 Associates with Nascent DNA. The amount of SMARCA5 on nascent chromatin following loss of HDAC1,2 function is shown. NIH3T3 cells were either treated with an HDAC1,2-selective inhibitor (898) or transfected with non-targeting (NT) or Hdac1,2 (H12) siRNA. Cells were labeled with BrdU following the above-mentioned treatments and ChIP with anti-SMARCA5 or rabbit IgG (negative control) was performed. Increasing volumes of ChIP DNA was spotted on the slot blot and the membrane was probed with anti-BrdU antibody. This figure is derived from our previous published work6. Please click here to view a larger version of this figure.
