Mesenchymal stromal cells (MSCs) are increasingly recognized for their therapeutic potential in a wide range of diseases, including lung diseases. Besides the use of bone marrow and umbilical cord MSCs for exogenous cell therapy, there is also increasing interest in the repair and regenerative potential of resident tissue MSCs. During development, including in the lung, the mesenchyme is an important source of developmental cues, and resident MSCs are a likely candidate to be at the center of this. Moreover, evidence is emerging that resident MSCs are perturbed in adult diseases, including cancer1,2 and fibrosis3. The main hurdle for the study of these resident tissue MSCs is the lack of a clear marker for the isolation and identification of these cells4. Stem cell antigen-1 (Sca-1) was identified in mice as a marker for a variety of tissue stem cells, and can be used for the isolation of L-MSCs5, but has unfortunately no known orthologs in other species6. Researchers have reported a variety of different isolation methods for the isolation of L-MSCs from either lung tissue or fluid. These vary from fluorescence activated cell sorting (FACS) based methods selecting for CD31–/CD45–/CD90+ cells7, CD31–/CD45–/epithelial cell adhesion molecule (EpCAM)-/Sca-1+ cells8, multidrug resistance transporter ATP binding cassette G (ABCG2) positive cells9 or Hoechst 33342 dye efflux10, to plastic adherence11,12 and migration out of minced tissue13.
The advantages of the herein presented method are several fold. By using a gentle enzymatic digestion and density gradient14, one obtains all the cells of the density range that include MSCs but exclude epithelial or endothelial cells. The subsequent plastic adherence step ensures that only the mesenchymal cells adhere and stay in culture, eliminating leukocytes. Most importantly however, the CD146+ selection step allows for the elimination of fibroblasts, as these cells do not express CD146. Expression of the cell adhesion molecule CD146 is positively correlated with multipotency, and is therefore a good marker to weed out fibroblasts from a mesenchymal cell population15-19. This is an advantage over using CD90 as a selection marker, as it is not only expressed in MSCs but also in lipofibroblasts5,20. In this protocol we have explicitly chosen a magnetic bead selection, as it is gentler on the cells, and the entire procedure can be done in sterile conditions. Another important advantage of this isolation method as opposed to the outgrowth method, is that it is relatively fast, 6-10 days as opposed to a month or more for the outgrowth method. Three to five days after the initial isolation the mesenchymal population is ready for CD146+ selection; after another three to five days the CD146+ cells are ready to be used for experiments or can be frozen for later use. The decreased time of culture improves the quality of the cells as MSCs transdifferentiate towards fibroblasts in prolonged ex vivo culture19. Lastly, because of the nature of the protocol, it is possible to apply this method to other species by simply choosing appropriate antibodies, or even to other organ systems by adjusting the choice of digestion enzymes and incubation time.
A detailed protocol of this isolation method is given below, and a schematic overview of the isolation and subsequent selection of the CD146+ subpopulation is provided in Figure 1A and 1B respectively. Additionally, details are included for passaging, freezing and thawing these cells.
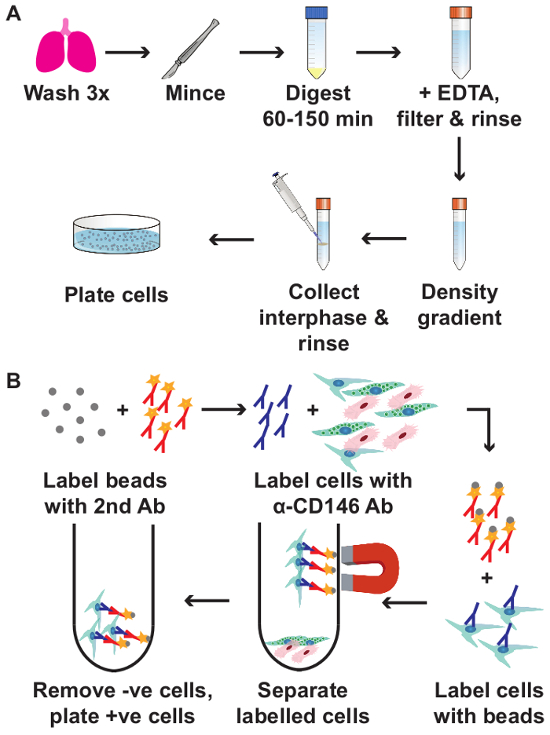
Figure 1. Schematic overview of the isolation of pulmonary mesenchymal cells (A) and subsequent CD146+ cell selection (B). min = minutes; EDTA = Ethylenediaminetetraacetic acid; 2nd Ab = secondary antibody; α-CD146 Ab = primary anti-CD146 antibody; -ve cells = CD146 negative cells; +ve cells = CD146 positive cells. Please click here to view a larger version of this figure.
Two of the most reliable physical characteristics of MSCs, density and plastic adherence, are used in the first part of this protocol to obtain the mesenchymal cell fraction of the lung that contains L-MSCs. Although the density gradient interphase will include monocytes and macrophages in addition to lung mesenchymal cells, the plastic adherence followed by 3-5 days of culture ensures that only the lung mesenchymal cells remain. Indeed this cell population expresses the classic MSC surface markers CD73, CD90 and CD146 and is negative for the markers CD34, CD45, major histocompatibility complex type II (MHCII)-RT1B, CD11b and CD79a, indicating that there are no longer any leukocytes present in the cell population (Figure 2A). Of interest is that out of the CD146+ subset, a range of 44.4-65.7% is also CD73+ and CD90+ (data not shown). Moreover, this cell population is capable of a colony forming unit (CFU) efficiency that at ~30% is much higher than generally reported for bone marrow MSCs or even other MSC types (Figure 2B), and differentiates along the three classic MSC lineages (Figure 2C).
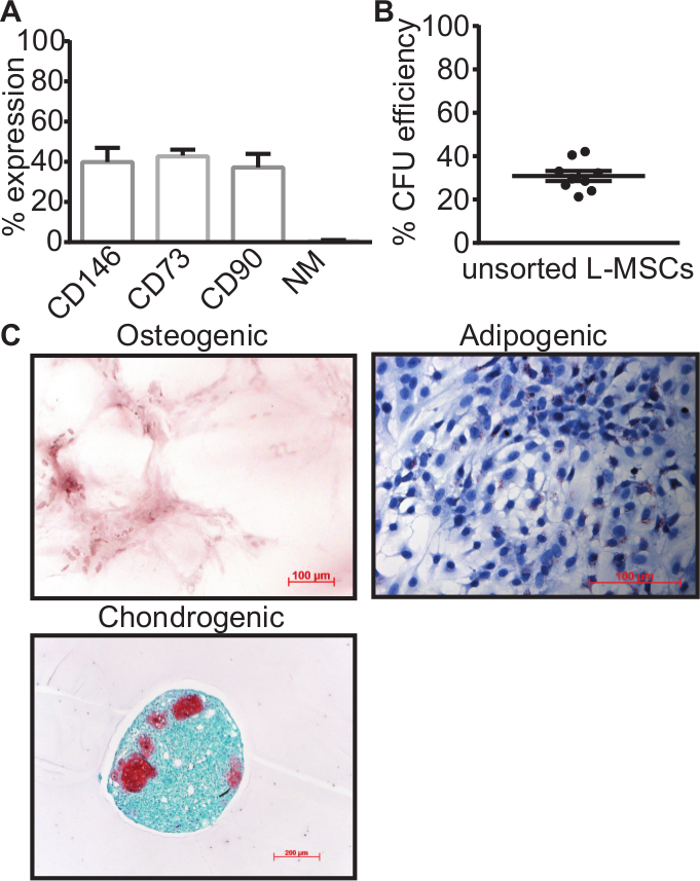
Figure 2. MSC characteristics after enzymatic digestion, density gradient separation and plastic adherence. (A) Expression of MSC-related surface markers CD146, CD73 and CD90 is present in part of the plastic adherent cell population, whereas the negative leukocyte markers CD11b, CD79a, CD34, CD45 and MHCII-RT1B were virtually not expressed. (B) Colony forming unit assay through limiting dilution assay (5 cells/cm2) indicated that ~30% of plastic adherent cells had a clonal capacity, indicative of MSCs. (C) Plastic adherent cells were capable of osteogenic matrix production (red), contained a higher number of small lipid vesicles (red) when induced with adipogenic differentiation medium compared to non-differentiated controls and formed chondrogenic like spheres containing chondrocytes (red). Data is presented as mean ± standard error of mean (SEM). All data were generated with passage 1-3 cells. NM = negative markers; CFU = colony forming units. Please click here to view a larger version of this figure.
However, the main caveat is that this population likely contains both L-MSCs and different subtypes of lung fibroblasts, the L-MSCs differentiated progeny23. Since multipotency is positively correlated with CD146 expression, and fibroblasts should have no expression of this marker, positive selection of the CD146+ subpopulation ostensibly resulted in a cell population that is highly enriched in L-MSCs. This is clearly demonstrated by a higher percentage of the cell population expressing MSC associated surface markers (Figure 3A), a considerably higher colony forming potential of ~80% (Figure 3B) and a stronger differentiation response in particularly the chondrogenic lineage (Figure 3C) compared to the total mesenchymal population that is obtained before CD146 selection. Of note is that these L-MSCs form only very few true adipocytes, but show many more spindle shaped cells that are filled with small lipid vesicles. These could reflect the lipofibroblast lineage that is crucial for both lung development and alveolar type II cell support. After CD146 selection, more adipocyte like cells filled with large lipid vesicles can be observed (Figure 3C) compared to before CD146 selection, yet the majority of cells are still the lipofibroblasts-like cells containing small lipid vesicles.
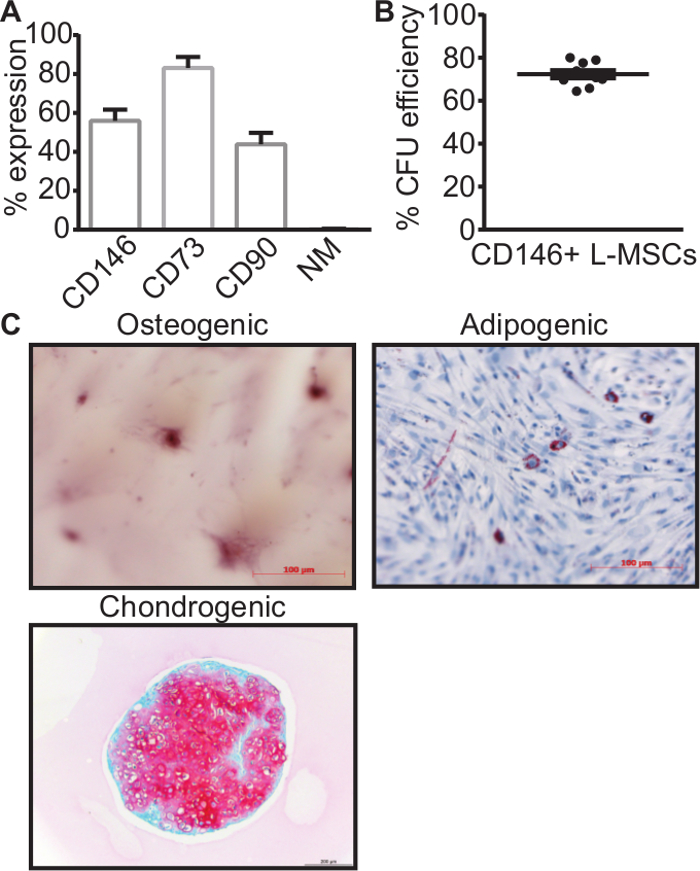
Figure 3. MSC characteristics after additional CD146+ magnetic bead selection. After positive selection of the CD146+ subpopulation, these cells showed a higher expression of MSC-related surface markers, CD73 in particular (A), a considerably higher CFU efficiency after single cell plating (B), and a stronger differentiation response for particularly the chondrogenic lineage and to a lesser extent the adipogenic lineage (C). Data is presented as mean ± SEM. All data were generated with passage 3 cells. NM = negative markers; CFU = colony forming units. Please click here to view a larger version of this figure.
