The grooming assay yields quantitative data to assess behavioral performance based on the relative remainder of accumulated dye left on the bodies of flies after a set time of measurement for grooming (30 min). Sample images of the sliding grooming chamber design and major steps of the assay are highlighted in Figure 1. Flies aggregate a significant amount of dye from immediate dusting by vortexing in the presence of dye (Figure 2d, 2e). Dusted flies can retain a range of dye accumulation post-assay (Figure 2f, 2g). Therefore, solubilizing dye accumulation in EtOH and measuring absorbance of individual samples provides a reproducible and highly quantitative assessment of grooming ability.
In this study of DopR in regulation of hindleg grooming, we compared the grooming performance of a strong hypomorph (Figure 3a, 3b) and null DopR mutant to a WT strain (Figure 3c, 3d). The DopR mutants retained significantly more dye than their WT or rescued counterparts, indicating that the DopR mutants were less efficient at grooming behavior (Figure 3a, 3c). To refine the understanding of the role of DopR, we performed a parallel experiment with rutabaga flies, carrying a strong hypomorphic mutation of calcium-dependent adenylate cyclase, which functions downstream of G-Protein Coupled Receptors in Drosophila. These mutants displayed higher dye accumulation levels than DopR mutants (Figure 3e, 3f). The data suggest a significant role for DopR in grooming behavior and a possible contribution by other GPCRs working in parallel or in different locomotor programs for grooming behaviors.
To investigate the differences between grooming abilities of foreleg vs hindleg grooming programs, we directly assessed dye accumulation on separable body parts during grooming (Figure 4). By dissecting the heads, wings, or "body" (abdomen/thorax/legs) of individual flies post-assay, we were able to reliably quantitate differences and similarities between genotypes. The findings supported an interpretation that no differences were present for foreleg grooming programs, as no significant differences were observed for heads of WT Vs. DopR mutants. However, significant differences were observed for both wing and body measurements between genotypes. This supplemental technique to perform the primary assay on body parts instead of whole flies allows a simple method to initially distinguish foreleg versus hindleg grooming programs. These results were further extended and supported by careful behavioral observations and video recording or tracking of individual components of the foreleg or hindleg behaviors. All results from Figures 2-4 are reproduced with permission from Genes, Brain, and Behavior15.

Figure 1: Drosophila Grooming Assay Workflow. This simplified flowchart outlines the major steps in the grooming protocol, highlighting some of the necessary materials and equipment. Please click here to view a larger version of this figure.

Figure 2: Quantification of Grooming based-on Dye Accumulation. (a) Grooming chamber. Individual flies and Brilliant Yellow dye are placed in individual wells (1 fly : 1 well). Dimensions and blueprints for production in supplemental data. (b, d, f) WT adult male Drosophila (+/+). (c, e, g) DopRf02676/DopRf02676 adult male Drosophila. (b, c) Flies before dusting. (d, e) Flies immediately after dusting by vortexing chamber, prior to grooming (time: 0 min). (f, g) Flies after grooming (time: 30 min). Scale bar = 400 µm. Reprinted with permission from Genes, Brain, and Behavior15. Please click here to view a larger version of this figure.
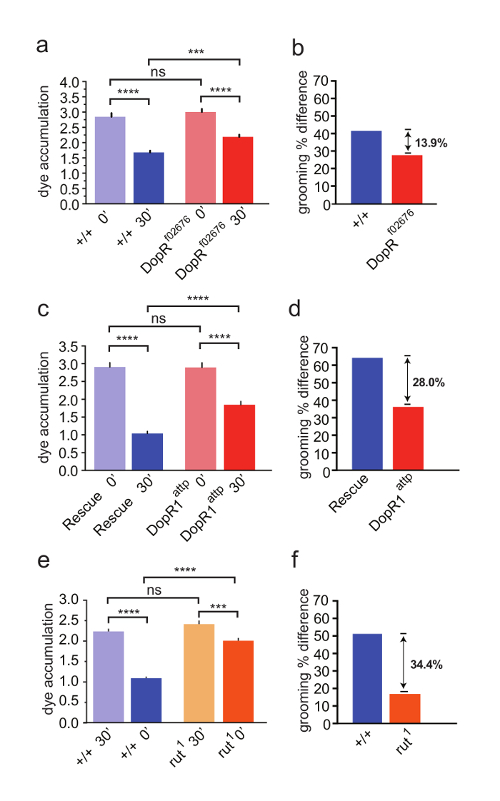
Figure 3: Dopamine Receptor (DopR/dDA1/dumb) is Required for Modulation of Grooming Behavior. (a, c, e) Grooming of wildtype (blue) and mutant flies (red/orange) measured at 0 min or 30 min after dusting. SEM is measured for each genotype and condition. (a, b) n = 43 flies per genotype and condition. (a) WT and DopRf02676 flies both exhibit grooming behavior (+/+ 30' compared to +/+ 0' = p value <0.0001, DopRf02676 30' compared to DopRf02676 0' = p value <0.0001). DopR flies fail to groom as well as WT animals (DopRf02676 30' compared to +/+ 30' = p value <0.001). Acute dusting of each genotype at 0' results in equivalent accumulation of dust (ns = not significant). (b, d, f) Grooming percent difference is calculated for each genotype (dye accum. at 0' – dye accum. at 30' / dye accum. 0' x 100) providing a relative value for comparing grooming behaviors. (c) DopRattp/DopRattp nullflies display a grooming deficit (DopRattp 30' compared to +/+ 30' p value <0.0001). (d) Grooming percent difference for DopRattp null. (c,d) n = 45 flies for each genotype or condition. e) rut1homozygous flies display a grooming deficit (rut1 30' compared to +/+ 30', p value = <0.0001). f) grooming index for rut1 flies. (e, f) n = 30 flies for each genotype or condition. Statistical analyses by One Way ANOVA and Bonferonni Correction. Reprinted with permission from Genes, Brain, and Behavior 15. Please click here to view a larger version of this figure.
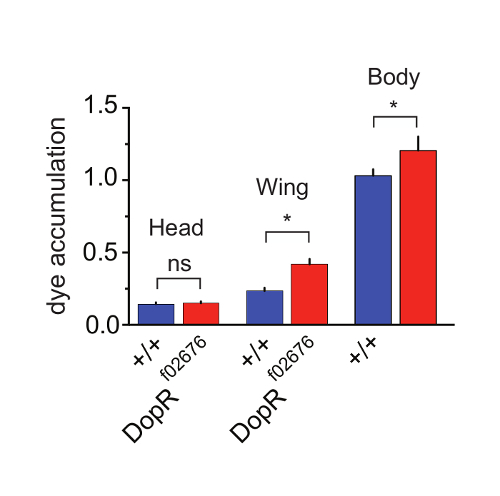
Figure 4: Dopamine Receptor Function Potentiates Hindleg Grooming. (a) Grooming of individual regions of WT (blue) and DopRf02676/DopRf02676 (red) measured at 30 min after dusting and subsequent dissection. p value for wing grooming = 0.0204. p value for body = 0.0302. n = 33-35 flies for all conditions. SEM is measured for each genotype and condition. Statistical analysis by One Way ANOVA and Bonferonni Correction. Reprinted with permission from Genes, Brain, and Behavior15. Please click here to view a larger version of this figure.
Supplemental Figure: Alternate Method for Visual Quantification of Grooming Behavior using NIH Image J Pixel Intensity Software. (A) Visualization of wildtype Drosophila under dissecting scope. Oval defines dorsal abdomen as the region of analysis for pixel intensity. (B) Visualization of dorsal abdomen after dusting with Ultra Green V10 fluorescent paint pigment. Standard filter for Green fluorescence captures the blue-green pigment. (C) WT animal after coating with UGV10 pigment (pregrooming). (E) WT animal after coating with UGV10 pigment (postgrooming). (D) DopRf02676 homozygous mutant after coating with UGV10 pigment (pregrooming). (F) DopRf02676 homozygous mutant after coating with UGV10 pigment (post-grooming). (G) Quantification of pixel intensity for all conditions. n = 15 flies for each genotype or condition. Statistical analyses by One Way ANOVA and Bonferonni Correction. ns = nonsignificant difference. *** represents a p <0.001. Please click here to download this file.
