Taking advantage of the PM-specific reporter mouse, we developed a multiplex immunostaining strategy to identify diverse endogenous myocytes as depicted in Figure 1. Following the reprogramming steps shown in Figure 2, induction of subtype-specific CMs can be detected as early as day 421, albeit at a low-rate. By day 14, the experiment can be stopped and assessed for sarcomere organization (Figure 3) and subtype-specification (Figure 4). Figure 5 summarizes the workflow of slide preparation for ICC (Figure 5 Panel A), and the quantification of iCLM subtype-specific cells (Figure 5 Panel B/C).
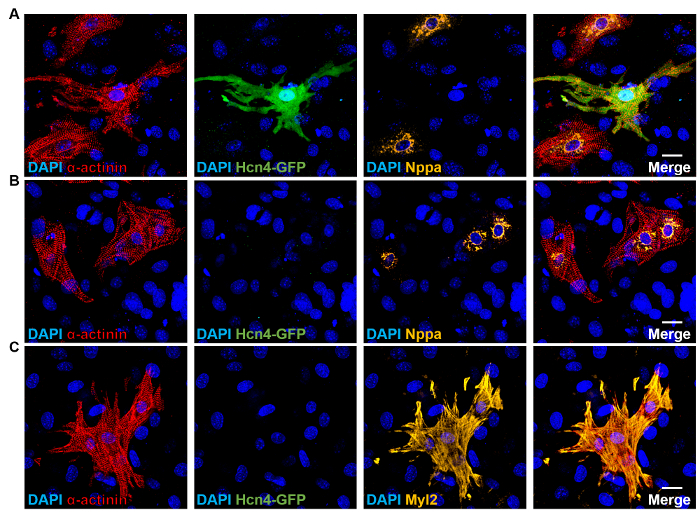
Figure 1: Subtype Diversity of Endogenous Cardiomyocytes. (A-B) Immunocytochemistry (ICC) staining of neonatal atrial cardiomyocytes from Hcn4-GFP reporter mice for α-actinin (sarcomere marker, red), Hcn4-GFP (PM marker, green), and Nppa (atrial marker, orange). (C) Immunocytochemistry staining of neonatal ventricular cardiomyocytes from Hcn4-GFP reporter mice for α-actinin (sarcomere marker, red), Hcn4-GFP (PM marker, green), and Myl2 (ventricular marker, orange). DAPI (blue): nuclear staining. Scale bars: 20 µm. Please click here to view a larger version of this figure.
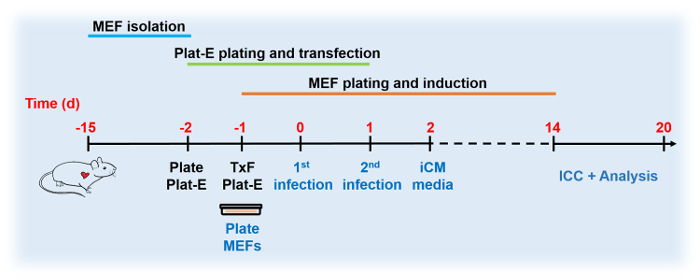
Figure 2: Reprogramming Timeline Schematic. Schematic representation of GHMT-induced Hcn4-GFP MEFs. The three major stages are depicted. Please click here to view a larger version of this figure.
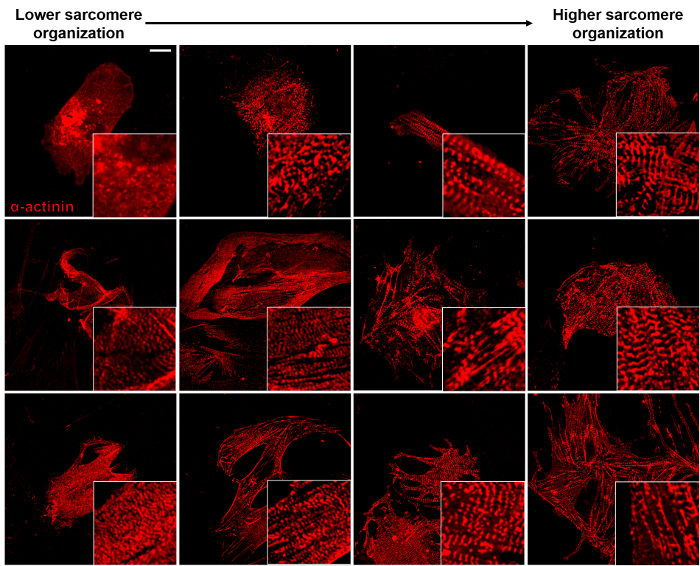
Figure 3: Degree of Sarcomere Organization. ICC staining of Hcn4-GFP MEFs 14 days after GHMT transduction for α-actinin (sarcomere marker, red) shows a diverse range of sarcomere organization. The degree of organization increases from left to right panels. Representative pictures of each level (n= 3). Scale bar: 20 µm. Please click here to view a larger version of this figure.
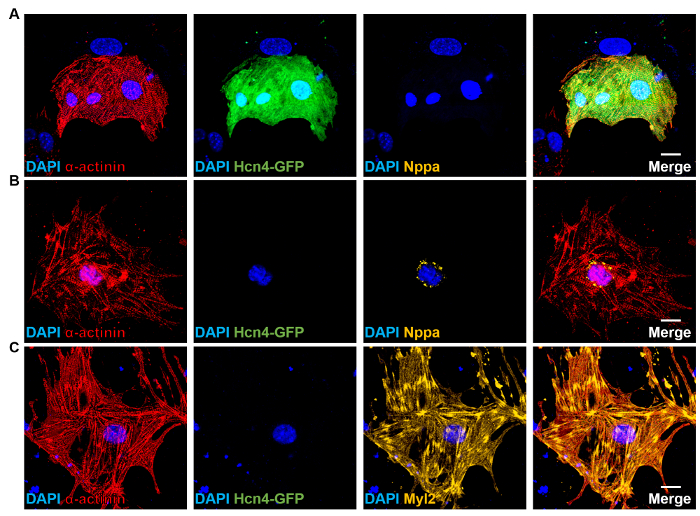
Figure 4: Subtype-specific Reprogrammed Cardiomyocytes. (A-C) ICC staining of GHMT-transduced Hcn4-GFP MEFs for α-actinin (sarcomere marker, red), Hcn4-GFP (PM marker, green), Nppa (atrial marker, orange), or Myl2 (ventricular marker, orange). DAPI (blue): nuclear staining. Scale bars: 20 µm. Please click here to view a larger version of this figure.
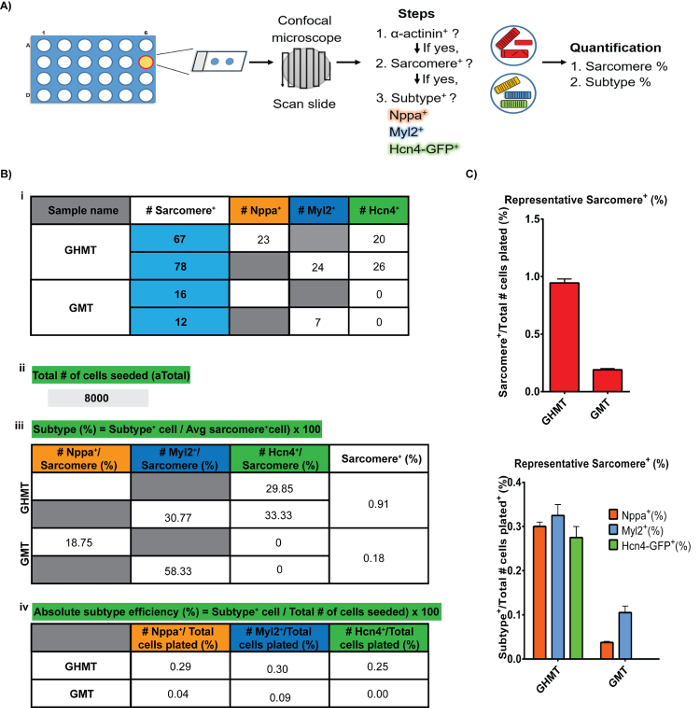
Figure 5: Image Acquisition and Analysis Workflow. Schematic representation for image analysis. Panel A depicts the priority order of assigning sarcomere+ and subtype-specificity to a cell. Panel B (i-iv) and C show the expected results from an average GHMT-iCLM experiment. Key points and formulas are shown in green. Please click here to view a larger version of this figure.
| iCLM media | ||
| Component | Volume (mL) | Final concentration |
| DMEM | 270 | |
| Medium 199 | 90 | |
| FBS | 50 | 10% |
| Insulin-Transferrin-Selenium G | 2.5 | 0.50% |
| MEM vitamin solution | 10 | 2% |
| MEM Amino Acids | 20 | 4% |
| Non-essential amino acids | 10 | 2% |
| Antibiotic-Antimycotics | 10 | 2% |
| B-27 supplement | 10 | 2% |
| Heat inactivated Horse Serum | 25 | 5% |
| Na-Pyruvate | 2.5 | 1.5 mM |
| Plat-E media (PE) | ||
| Component | Volume (mL) | Final concentration |
| DMEM | 450 | |
| FBS | 50 | 10% |
| Penicillin/Streptomycin | 5 | 1% |
| Puromycin | 0.05 | 1 μg/mL |
| Blasticidin | 0.5 | 10 μg/mL |
| Fibroblast medium (FB) | ||
| Component | Volume (mL) | Final concentration |
| DMEM | 450 | |
| FBS | 50 | 10% |
| Penicillin/Streptomycin | 5 | 1% |
| Glutamax | 5 | 1% |
| Transfection medium (TxF) – Filtered (0.45 μm) | ||
| Component | Volume (mL) | Final concentration |
| DMEM | 450 | |
| FBS | 50 | 10% |
| Immunocytochemistry (ICC) staining buffer | ||
| Component | Volume (mL) | Final concentration |
| 1x PBS | 5 | |
| 1x Universal blocking buffer | 5 | |
Table 1: Culture Medium. Table summary for the preparation of the several mediums used during GHMT-induced reprogramming.
| A) Cell seeding and transfection | ||||||
| Plate/Dish | Surface Area (cm2) | Seeding density (cells) | Growth medium (mL) | Total DNA amount to transfect (μg) | Transfection Reagent (μL) | Reduced Serum Media (μL) |
| 15 cm plate | 152 | 1.00E+06 | 20 | 25 | 75 | 600 |
| 10 cm plate | 55 | 5.50E+06 | 10 | 9 | 27 | 300 |
| 6 cm plate | 21 | 2.20E+06 | 4 | 3.5 | 10.5 | 105 |
| 6 well/x1 | 9 | 1.00E+06 | 2 | 2 | 6 | 60 |
| 12 well/x1 | 4 | 4.00E+05 | 1 | 0.5 | 1.5 | 15 |
| 24 well/x1 | 2 | 2.00E+05 | 0.5 | 0.3 | 0.9 | 9 |
| 48 well/x1 | 1 | 1.70E+05 | 0.25 | 0.15 | 0.45 | 4.5 |
| B) Fibroblast seeding and induction | ||||||
| Plate/Dish | Fibroblast seeding density (millions) | Approximate infection units for iCLM | ||||
| 6 cm plate | 0.22-0.33 | 5.00E+7 (~ 5 mL) | ||||
| 6 well/x1 | 0.1-0.15 | 3.00E+7 (~ 3 mL) | ||||
| 12 well/x1 | 0.04-0.06 | 1.30E+7 (~ 1 mL) | ||||
| 24 well/x1 | 0.02-0.03 | 6.50E+6 (~ 0.8 mL) | ||||
| 48 well | 0.001-0.015 | 3.00E+6 (~ 0.4 mL) | ||||
Table 2: Seeding, Transfection and Induction Formats. (A) Table summary for the plating and transfection of cells. (B) Seeding density and approximate infection units (or viral supernatant) needed to induce MEFs into cardiomyocyte-like cells.
