HPLC chromatograms of the fluorescent labeled Neu5Ac and the Neu5Gc standards are depicted in Figure 2. Using the herein described method, DMB-labeled Neu5Gc typically elutes between 7 – 9 min elution time, and DMB-Neu5Ac between 10 – 12 min. Several smaller peaks in the chromatogram usually appear between 2 – 6 min. These peaks represent unreacted DMB and reaction intermediates25.
Figure 3 shows representative chromatograms of cell lysates. Compared to the chromatograms of the DMB-labeled standards (Figure 2), in these chromatograms several undefined peaks are visible. This is most likely due to the internal impurity of cell lysates and to the fact that DMB not only reacts with sialic acid species, but also with other α-keto acids present in cell lysates, including pyruvate, succinate, and α-ketoglutarate26. The presence of small amounts of sialic acids bearing O-acetyl modifications that have not been cleaved by acetic hydrolysis might also be discussed as a reason for these undefined peaks23. Chromatograms from lysed untreated cells are compared with chromatograms from cells that have previously been treated with ManNAc or its analogs. As shown here, treatment with ManNAc often leads to an almost entire depletion of Neu5Gc on the cell surface. In chromatograms of lysed cells treated with Neu5Prop or Neu5But, peaks are visible that do not appear in chromatograms of untreated cells. These peaks (for ManNProp treated cells at approximately 19 min elution time and for ManNBut treated cells at 42 min) indicate the appearance of the corresponding non-natural sialic acids, Neu5Prop and Neu5Gc. As described in section 8.3 of the protocol, the HPLC chromatograms can be analyzed in order to quantify the amounts of the different sialic acid species in the cell lysates.
The fact that distinct HPLC retention peaks correspond to certain sialic acid species was verified via mass spectrometry. Representative ESI-MS data from collected HPLC peaks of interest are depicted in Figure 4: the spectrum of the collected DMB-Neu5Ac retention peak in the upper left panel, DMB-Neu5Gc in the upper right panel, and the two non-natural sialic acid species, DMB-Neu5Prop and DMB-Neu5But in the lower left and right panels. The reaction with DMB leads to an increase in the molecular mass of 116.2 Da, compared to the sialic acid species that are not labeled with this dye. In the mass spectra, when displaying the positive mass/charge ratio of the injected samples between 300 and 700, several peaks are visible. This is due to adduct formation of the respected DMB-labeled probe. Besides the protonated ion [M+H]+, the sodium adducts [M+Na]+ and [M+2Na-H]+ appear. As acetonitrile (ACN) is present during the LC analysis, it may be found as an adduct in the mass spectra (shown for DMB-Neu5Gc, Figure 4: upper right panel). Due to fragmentation caused by collisional decompensation activations generated in the electronspray transport region between the capillary and the first skimmer, dehydrated sialic acid species [M+H]+-H2O can also be observed (shown for DMB-Neu5Ac, Figure 4: upper left panel)23,27. In the LC setup, DMB-Neu5Gc elutes shortly after the unreacted or partly reacted DMB-species. Therefore, the collected DMB-Neu5Gc probe may contain impurities caused by these reaction intermediates. This serves as an explanation for the appearance of undefined peaks in the mass spectrum of DMB-Neu5Gc (Figure 4: upper right panel).
Treatment of Kelly neuroblastoma cells with ManNProp or ManNBut leads to reduced expression of polysialic acid on NCAM, as shown by Western blot analysis of the cell membrane from lysed cells (Figure 5). Polysialic acid normally appears as a smear do to its heterogeneity in size and charge of (≈ 200 kDa). Treatment with ManNAc analogs leads to the reduction of polysialic acid on the cells surface: the longer the aliphatic N-acyl side chain, the stronger the reduction28.
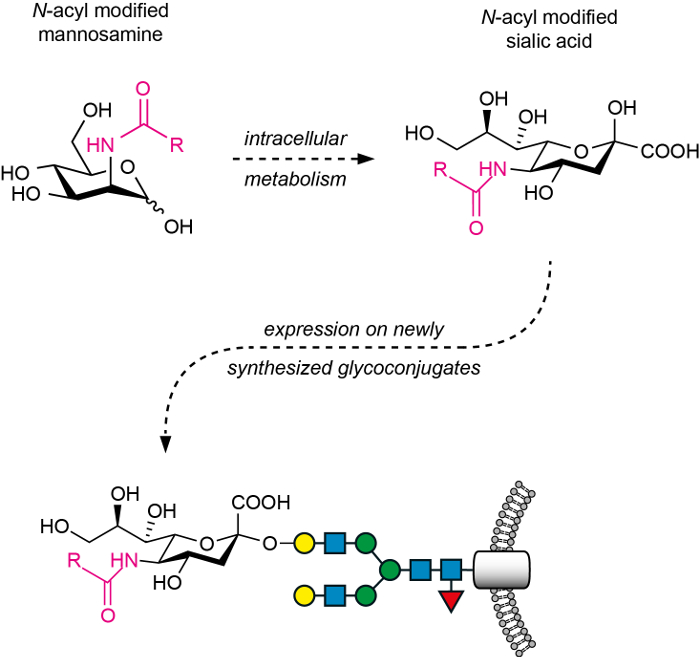
Figure 1: The general principle of MGE with N-acyl modified mannosamines. After treatment with the N-acyl modified mannosamines, these ManNAc analogs are unidirectionally metabolized (intracellular metabolism) to the corresponding non-natural sialic acids. These are transported to the Golgi, transferred to the respective oligosaccharide acceptor by sialyltransferases, and expressed on the cell surface as sialosides (here, a glycoprotein). Please click here to view a larger version of this figure.
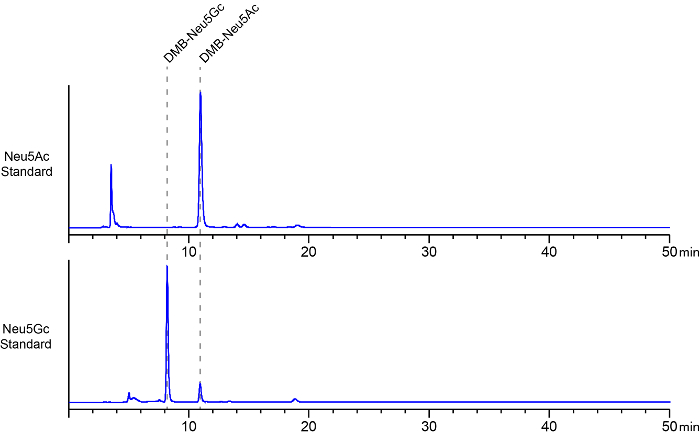
Figure 2: Representative chromatograms of DMB-labeled sialic acid standards. DMB-labeled Neu5Ac (upper panel) and Neu5Gc (lower panel) were analyzed by HPLC. Retention times (dashed lines) and peak areas of both sialic acids were determined after injecting 10 ng of the respected DMB-labeled species into the HPLC system. In the herein shown representative chromatograms, DMB-Neu5Gc eluted after approximately an 8 min retention time and DMB-Neu5Ac at 11 min, respectively. Several smaller peaks in the chromatogram usually appear between 2 – 6 min, representing unreacted DMB and reaction intermediates. A small fraction of DMB-labeled Neu5Ac appears as a minor impurity in the DMB-Neu5Gc standard (lower panel). This figure is modified from reference22. Please click here to view a larger version of this figure.
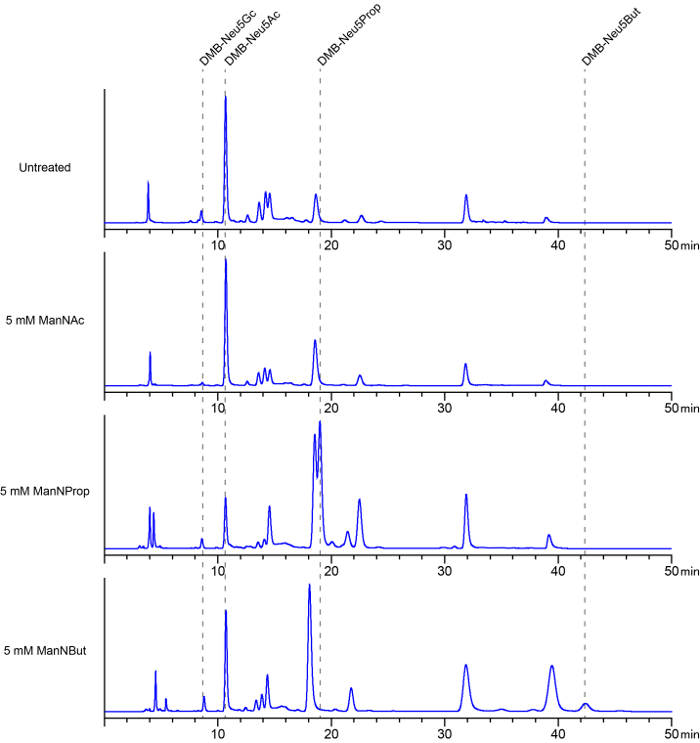
Figure 3: Characterization of membrane-bound sialic acids by DMB-HPLC. Cells were cultured in the absence (untreated, upper panel) or presence with 5 mM ManNAc (second panel from top), ManNProp (third panel from top), or ManNBut (lower panel), respectively for 7 days. The medium was changed every 24 h. Cells were harvested and membrane fractions were prepared, labeled with DMB, and analyzed by HPLC. By comparison with the chromatograms of DMB-labeled sialic acid standards (Figure 2), peaks at the retention times between 8-9 min were identified as DMB-Neu5Gc, and peaks between 10 – 11 min as DMB-Neu5Ac. Compared to the peaks obtained from injecting DMB-labeled sialic acid standards, chromatograms from cell lysates show additional, undefined peaks. This is due to the internal impurity of the probes, as well as to the fact that DMB can react not only with sialic acid present in the cell lysates, but also with other α-keto acids present in cell lysates, including pyruvate, succinate, and α-ketoglutarate. Peaks that only appear in the chromatograms of samples from cells cultured in the presence of N-acyl modified mannosamine analogs indicate the corresponding DMB-labeled non-natural sialic acids. This figure is modified from reference22. Please click here to view a larger version of this figure.
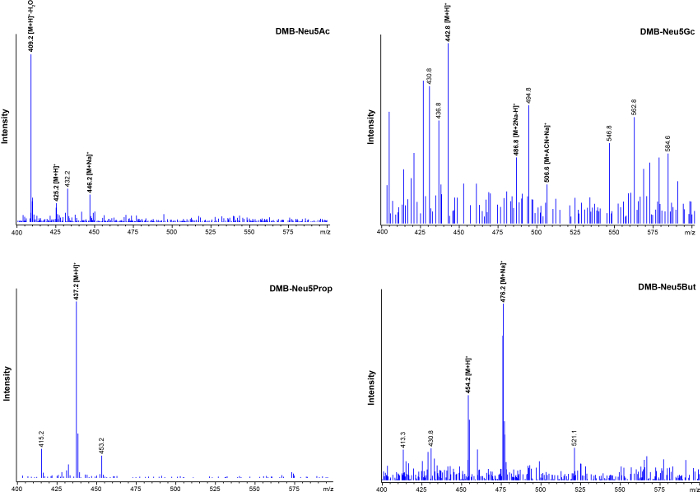
Figure 4: Mass spectra of DMB-labeled sialic acid species found in cell lysates. HPLC retention peaks of interest (see Figure 3) were collected and subsequently analyzed by ESI-MS. 20 µL of the collected retentions peaks were injected into the LC-MSD system. Reaction with DMB leads to an increase in the molecular mass of 116.2 Da compared to the corresponding unreacted sialic acid species. The positive mass/charge ratio spectrograms obtained from different sialic acid species are depicted (DMB-Neu5Ac: upper left panel, DMB-Neu5Gc: upper right panel, DMB-Neu5Prop: lower left panel, DMB-Neu5But: lower right panel). Due to adduct formation several peaks are visible. Besides the protonated ion [M+H]+, sodium adducts [M+Na]+ and [M+2Na-H]+ appear. Acetonitrile, which is present during the LC analysis, can also be found as an adduct (upper right panel). Dehydrated DMB-Neu5Ac, which is generated by fragmentation in the instrument [M+H]+-H2O can be observed (upper left panel). The collected DMB-Neu5Gc probe may contain impurities caused by unreacted DMB, as well as reaction intermediates seen as additional undefined peaks (upper right panel). ACN, acetonitrile; H, hydrogen; Na, sodium. This figure is modified from reference22. Please click here to view a larger version of this figure.
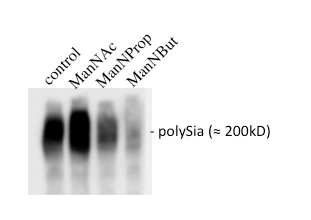
Figure 5: Analysis of the polysialylation of NCAM. Kelly neuroblastoma cells were cultured in the absence or presence of 5 mM ManNAc, ManNProp, or ManNBut for 7 days. Cells were harvested and proteins were separated on 8% SDS-acrylamide gels and analyzed by Western blot using monoclonal anti-polySia antibody 735. Note that polySia appears as a smear due to its heterogeneity in number of Sia, resulting in different sizes and charges of polySia.
