Our technique was successfully applied to determine the relative efficacy of the trans (Figure 5A) versus cis (Figure 5B) isomers of palmitoleic acid. Our representative data demonstrates that trans-palmitoleic acid is a more effective ligand for Or47b ORNs when compared to the cis isoform (Figure 5C). A single neuron was recorded from each fly, with twelve flies recorded per dosage curve, for a total of 24 flies. The collective data were obtained from three independent repeats of the experiments, with 8 flies recorded in each. The error bars represent the s.e.m.
Of note, the distance between the opening of the odor cartridge and the head of the fly has a significant influence on the outcome of the recording. To elicit a significant response to palmitoleic acid in Or47b ORNs, we presented the odorant at close range, around 4 mm away from the antenna1 (Figure 6A). When palmitoleic acid is presented further away from the antenna (~11 mm), we could hardly observe any significant response from the same Or47b ORNs (Figure 6B). These results highlight the importance of the close-range presentation of palmitoleic acid (Figure 6C-D). The data were collected from parallel experiments from 6 male flies (Berlin, 7 d old). A single Or47b ORN was recorded/fly. The error bars represent the s.e.m.

Figure 1: Cartridge and Olfactometer Setup. (A) Preparation of odor cartridges. From left to right: a standard 200-µL pipette tip, the first and second cartridge sections, and a completed odorant cartridge. (B) The cartridge connected to the olfactometer, showing the downward angling of the second section. (C) Olfactometer setup depicting the odor delivery tube mounted on the micromanipulator, with an attached odorant cartridge. Please click here to view a larger version of this figure.

Figure 2: Drosophila Preparation. (A) A complete preparation, showing the relative positions of the fly, coverslip, and holding rod. (B) Close-up view of the prep, showing the positioning of the fly, its antennal orientation, and its clypeus. The holding rod is placed over the arista, securing the third antennal segment to the double-sided tape. (C) Rig setup. All major components are annotated. Please click here to view a larger version of this figure.
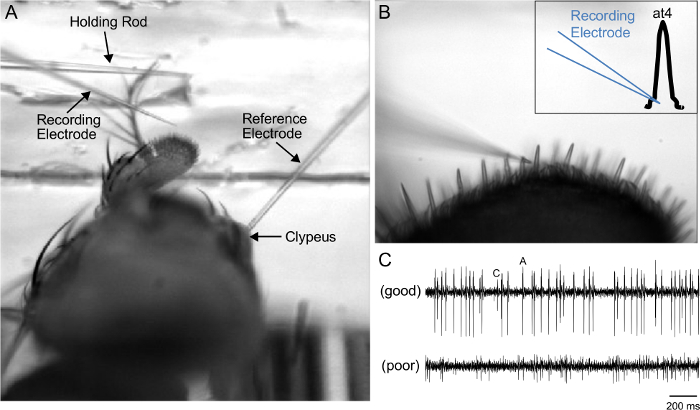
Figure 3: Identification of the at4 Sensillum for SSR. (A) 4X view of the prep, showing the reference electrode inserted in the clypeus, the holding rod atop the arista, and the recording electrode positioned near the third antennal segment. (B) 50X view of the electrode, poised for insertion into the at4 trichoid. Inset: Illustration of the position of the recording electrode. (C) Representative SSR traces of baseline spike activity, demonstrating good (top) or poor (bottom) signal-to-noise ratio. Good signal-to-noise ratio permits the reliable identification of at4A and at4C spikes. Please click here to view a larger version of this figure.

Figure 4: Cartridge Placement. (A) The odorant cartridge is aimed squarely at the head of the fly from a distance of a few mm. (B) Another view of the prep and olfactometer from a different angle. (C) A close-up view of the prep and olfactometer, showing the position of the odorant cartridge above the fly-prep slide. Please click here to view a larger version of this figure.
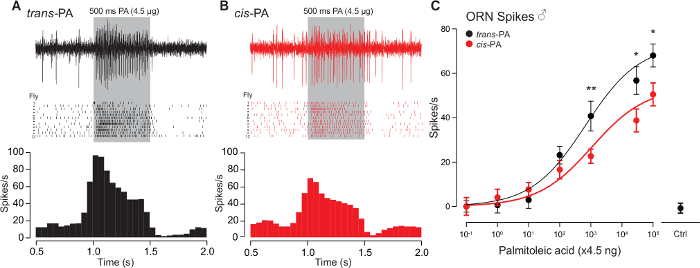
Figure 5: Representative Traces and Dosage Curves of Or47b ORNs in Response to cis– or trans-palmitoleic Acid. (A-B) SSR from the at4A ORNs that express the Or47b receptor with trans– (A) or cis-palmitoleic acid (B). Recordings were performed with 7-day-old WT Berlin males. Corresponding spike rasters (middle) and a peri-stimulus time histogram (bottom, binned at 50 ms) are shown below the sample traces (n = 12). (C) Dose-response curves comparing the Or47b ORN spike responses to cis– or trans-palmitoleic acid. Mean ±s.e.m. (*p <0.05; **p <0.01; t-test). Ctrl: Negative control without palmitoleic acid. Please click here to view a larger version of this figure.
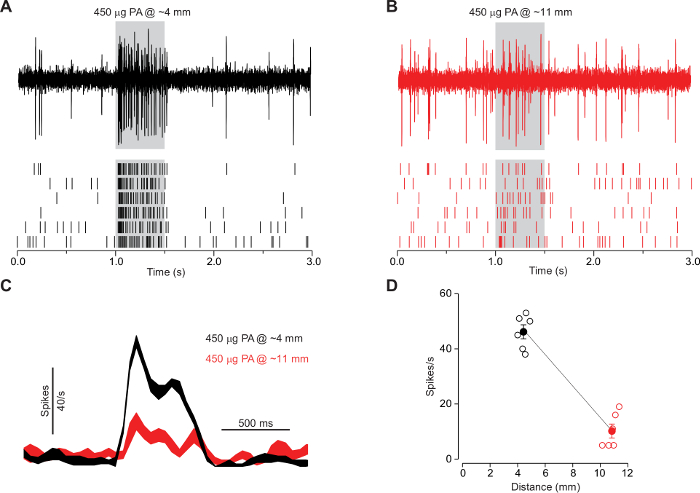
Figure 6: The activation of at4A by palmitoleic acid requires close-range stimulation. (A-B) SSR from the at4A ORNs in 7-day-old wildtype Berlin males. cis-palmitoleic acid was delivered at a close range (~4 mm) or further away (~11 mm) (n = 6). (C) Comparison of the corresponding spike responses (binned at 50 ms, smoothed peri-stimulus time histograms). (D) Comparison of the corresponding average spike responses. The responses of at4A to palmitoleic acid drop markedly as the stimulus distance increases. Reprinted with permission from Figure S4 in reference1. Please click here to view a larger version of this figure.
