We administered bioluminescence-labeled EHEC (~ 109 bacterial cells) to 6-week old female C57BL/6 mice by oral gavage. After oral inoculation of EHEC to mice within 1 h, the animals were examined for bioluminescent signal by the in vivo imaging system as shown in Figure 7. The results showed a strong bioluminescent signal in gavage mice with bioluminescence-labeled EHEC. We examined the signals on 2 days post infection. As shown in Figure 8A, the mice inoculated with bioluminescence-labeled wild-type EHEC EDL933 showed intense bioluminescent signals even after 2 days post infection, which suggested EHEC colonized in hosts by 2 days. We also intragastrically infected bioluminescence-labeled EDL933ΔrfaD (ΔrfaD) to mice (Figure 8A). This mutant, defected in lipopolysaccharide (LPS), has been shown to reduce colonization in the host in our previous study. As shown in Figure 8A, there is no bioluminescent signal detected in ΔrfaD-infected mice, which suggests that there are no or less bacteria cells colonized in the mice. Quantification of the fluorescent signal is shown in Figure 8B. Next, the location of these bioluminescence-labeled bacteria was determined. The infected mice were sacrificed humanely and their whole intestine removed. The intestines of mice 2 days post infection were positioned on 9 cm Petri dishes and imaged ex vivo (Figure 9A). The intestinal tissues of bioluminescence-labeled EDL933 infected mice revealed a significant increase in bioluminescent signals in the cecum and colon, which suggest that these bioluminescent EHEC colonized in the cecum and colon of infected mice for 2 days at least. In contrast, mice infected with bioluminescence-labeled ΔrfaD (Figure 9A), revealed decreased bioluminescent signal in their intestinal tissue, which is consistent with the in vivo image (Figure 8A). Quantification of the fluorescent signal is shown in Figure 9B.
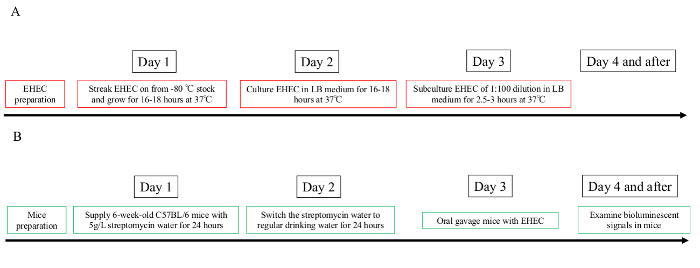
Figure 1: Timeline of the experimental preparation flow chart.
Overview of the timing needed to prepare bioluminescent EHEC bacteria and pretreat mice with streptomycin. (A) EHEC preparation. (B) Mice preparation. Please click here to view a larger version of this figure.
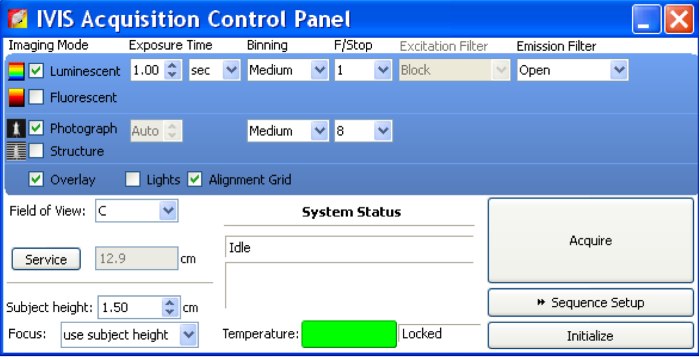
Figure 2: In vivo imaging system acquisition control panel.
Before imaging samples, open IVIS Acquisition Control Panel. Select "Luminescent," "Photograph," and "Overlay." Set Exposure Time as "Auto." Set Binning as "Medium." Set ƒ/stop as 1 for luminescent and 8 for photograph. ƒ/stop controls the amount of light received by the CCD detector. Once samples are ready for imaging, click "Acquire" to acquire images. Please click here to view a larger version of this figure.
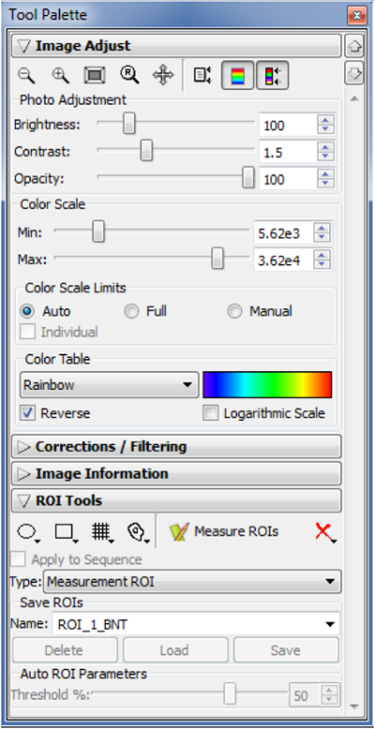
Figure 3: Tool Palette panel.
After image acquiring, use the Tool Palette panel for quantifying bioluminescent intensity. Open the Tool Palette panel and image the data. Choose one of the ROI Tools to range the bioluminescent signals on images. Please click here to view a larger version of this figure.
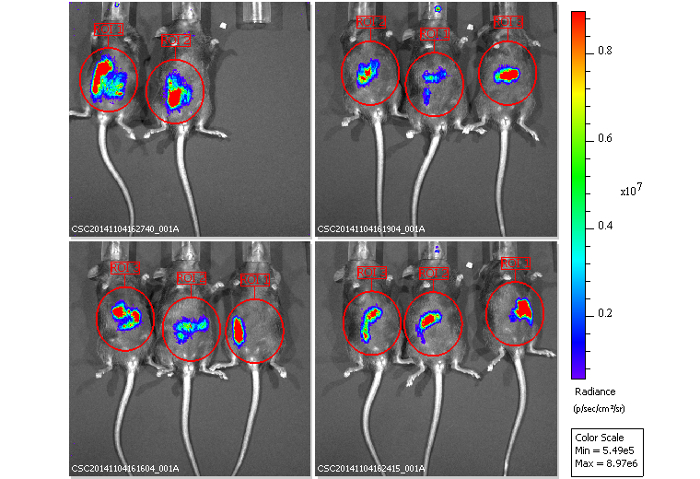
Figure 4: Bioluminescent signal from sample for quantification.
Bioluminescent signal area on images encircled by ROI Tools. All bioluminescent signals shown here are in the red circle. Please click here to view a larger version of this figure.
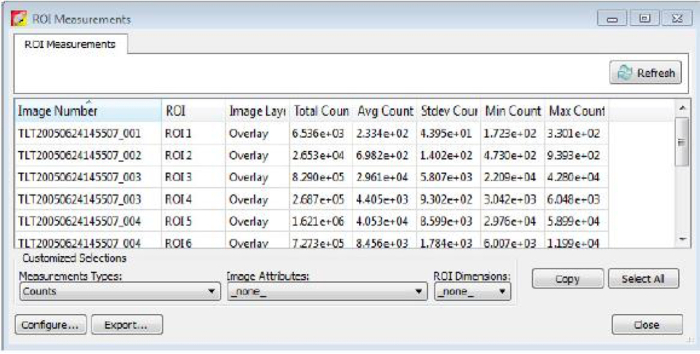
Figure 5: ROI measurements.
After circling bioluminescent signals and clicking "Measure ROIs" on the Tool Palette panel, values are presented as shown. The values of the column Total Flux (p/s) are used for the bioluminescent intensity quantification. Please click here to view a larger version of this figure.
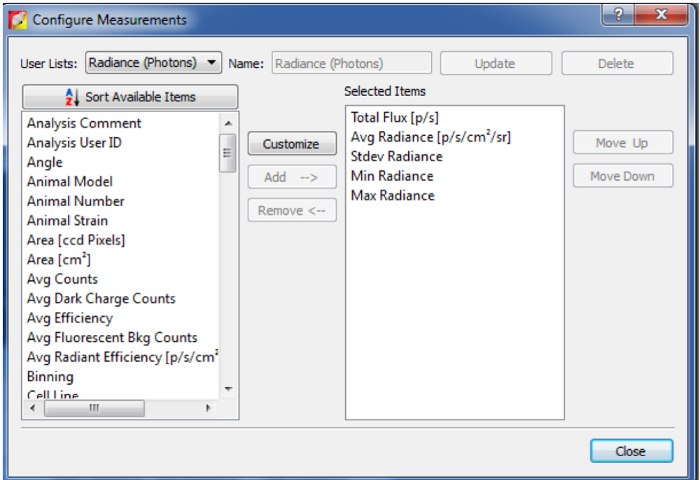
Figure 6: Add different quantification information.
By clicking on Configure Measurement on the left corner of the ROI Measurements panel, you can select other desired quantification values/information. Please click here to view a larger version of this figure.
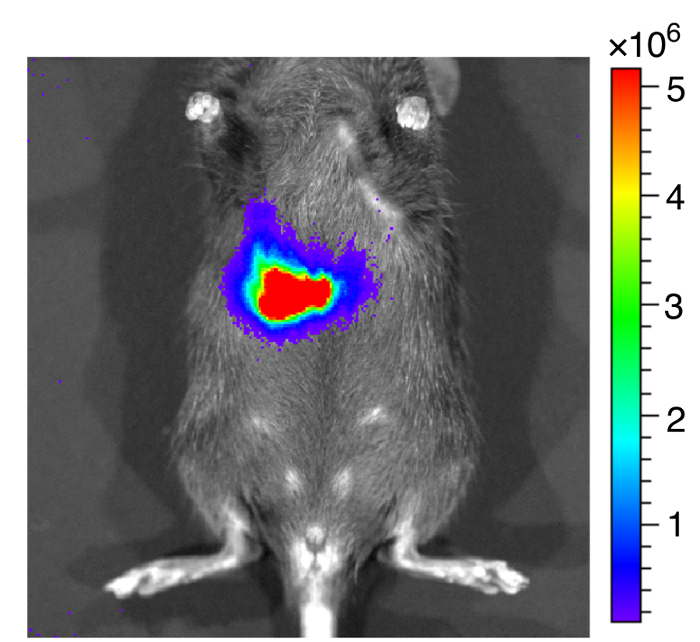
Figure 7: Representative image of mice after inoculated with bioluminescent EHEC.
Representative image of mice inoculated with bioluminescent EHEC by oral gavage within 1 h. The color scale represents the radiance (p/s/cm2/sr). Please click here to view a larger version of this figure.
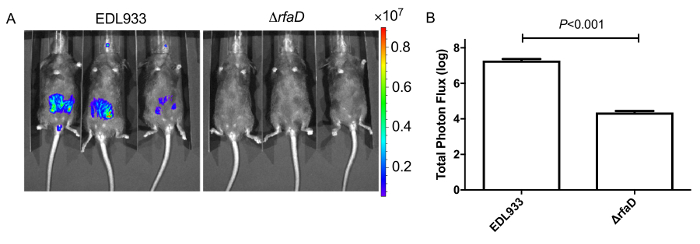
Figure 8: Images of mice inoculated with bioluminescence-labeled EHEC after 2 days.
(A) Represent image of mice inoculated with bioluminescent wild-type EHEC EDL933 and EDL933:ΔrfaD by oral gavage after 2 days post infection. (B) Quantification of bioluminescence intensity of mice infected with EHEC. Error bars indicate the standard deviations. Representative images are shown.All experiments were conducted independently three times with 2 – 3 animals each time, and error bars indicate the standard deviations. P-values denote the results of statistical analysis by t-test. The color scale represents the radiance (p/s/cm2/sr). Please click here to view a larger version of this figure.
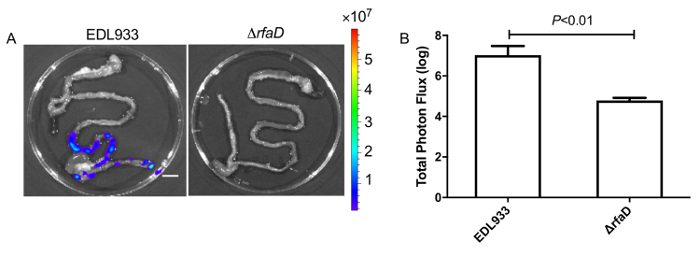
Figure 9: Images of intestinal tissues of infected mice with bioluminescence-labeled EHEC.
(A) 2 days after inoculation with bioluminescence-labeled EHEC, the mice were euthanized and whole intestinal tissues were removed and imaged ex vivo. Representative images are shown.(B) Quantification of bioluminescence intensity of intestinal tissues from mice infected with EHEC. All experiments were conducted independently three times with 2 - 3 animals each time, and error bars indicate the standard deviations. P-values denote the results of statistical analysis by t-test. The color scale represents the radiance (p/s/cm2/sr). Scale bar represents 1 cm. Please click here to view a larger version of this figure.
| Steps | Temperature | Time | Number of cycles |
| Initial denaturation | 95 °C | 10 min | 1 |
| Denaturation | 95 °C | 30 sec | 35 |
| Annealing | 58.4 °C | 30 sec | |
| Extension | 72 °C | 1.5 min | |
| Final extension | 72 °C | 10 min | 1 |
| Hold | 4 °C | ∞ | 1 |
Table 1: Polymerase chain reaction (PCR) conditions
| Primers name | Sequence |
| nptII F | 5’CCTATGCATAATAATTCCGCTAGCTTCACG3’ |
| nptII R | 5’GCTCCACCGATAATATTCCTGAGTCATACT3’ |
Table 2: Primer sequences used to amplify nptII
