The primary result of interest is the difference in accuracy between trials with and without an ISI. In particular, the operational definition of change blindness in the flicker paradigm is significantly reduced change-detection accuracy on trials with an ISI relative to trials without an ISI. This effect can be seen in Figure 3, which shows previously published data29. In that experiment, pigeons detected changes to stimuli consisting of color elements (the type depicted on the left side of Figure 1). As shown in the figure, pigeons saw 10 kinds of stimuli, differing based on the number of change repetitions (2, 4, 8, 16, or 32 on the x-axis) and presence of a 250 ms ISI (separate lines). Average accuracy on trials with an ISI (M = 76.4%) was worse than on trials without an ISI (M = 79.6%), F(1, 3) = 11.338, p = .043, partial η2 = .791. This figure also shows that this difference in accuracy indicative of change blindness was present for each number of repetitions tested.
Figure 3 also shows a secondary result of interest: change detection accuracy is influenced by the number of repetitions presented. In particular, change detection accuracy increased along with the number of repetitions, F(4, 12) = 11.104, p = .001 partial η2 = .787. This pattern is consistent with the interpretation that pigeons engaged in a serial search, much the way humans do in similar change detection tasks9, and supports the possibility that change detection requires an active search process requiring attention. Additional repetitions afford additional opportunities to consider more potential change locations, and attention gets directed to different spatial locations accordingly. Lower accuracy on trials with few repetitions may largely be a consequence of not having sufficient time to consider the actual change location before stimulus presentation was complete.
Figure 4 summarizes in a parallel fashion additional previously published results from the same lab30. The experiment presented pigeons with stimuli consisting of line segments (the type depicted in the right side of Figure 1), again for a fixed number of repetitions (1, 2, 4, 8, or 16) and with or without an ISI. Note that the condition depicted in the bottom panel shows both of the effects described above in Figure 3. Accuracy on trials with an ISI (M = 45.6%) was worse than on trials with no ISI (M = 75.8%), F(1, 3) = 60.571, p = .004 partial η2 = .953. Furthermore, accuracy increased along with the number of repetitions, F(1, 3) = 23.452, p < .001 partial η2 = .887. Thus, the same pattern of findings relevant to change blindness were obtained using the same general procedure, but using different kinds of stimulus elements.
The top panel of Figure 4 shows results from a different condition of the same experiment, in which the timing of change displays was manipulated. This condition revealed no change blindness effect. That is, there was no difference in accuracy between trials with (M = 66.1%) and without (M = 67.4%) an ISI, F(1, 3) = 0.189, p = .693 partial η2 = .059. This negative result suggests that change blindness can be dependent on the particular details of stimulus presentation, and that investigation of factors such as timing and salience may be informative in the future.
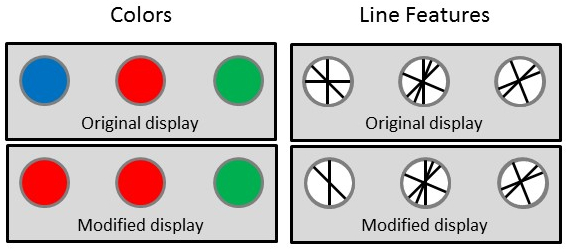
Figure 1: Examples of paired (original and modified) stimulus displays that make up change detection displays. Displays on the left consist of color elements (representative of displays from Herbranson & Jeffers, 201729). Displays on the right consist of line segments (representative of displays from Herbranson & Davis, 201630). In both examples, the change is on the leftmost key.
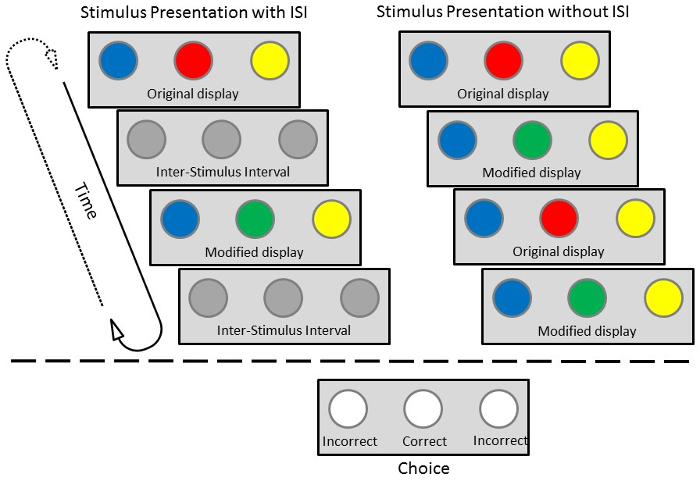
Figure 2: Schematic depiction of a trial with (left) or without (right) an ISI. The top section represents the stimulus presentation portion of a trial. The sequence on the left includes an ISI between each consecutive display. The sequence on the right does not. In both sequences, the change is on the center key (red-green). The bottom panel illustrates the subsequent choice portion of a trial. The correct response is the center key, corresponding to the location of the change. Please click here to view a larger version of this figure.
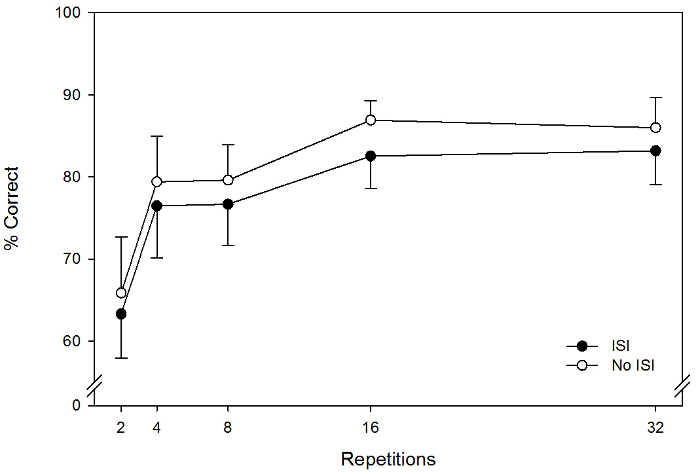
Figure 3: Rates of accurate change detection from Experiment 1 of Herbranson and Jeffers, 201729. This experiment used 250 ms display times and ISIs of either 250 ms or 0 ms). Error bars represent one standard error. The operational definition of change blindness is worse change detection on trials with an ISI (filled circles) relative to trials without an ISI (open circles). Note also that accuracy improves with the number of repetitions, indicating a serial search. Please click here to view a larger version of this figure.
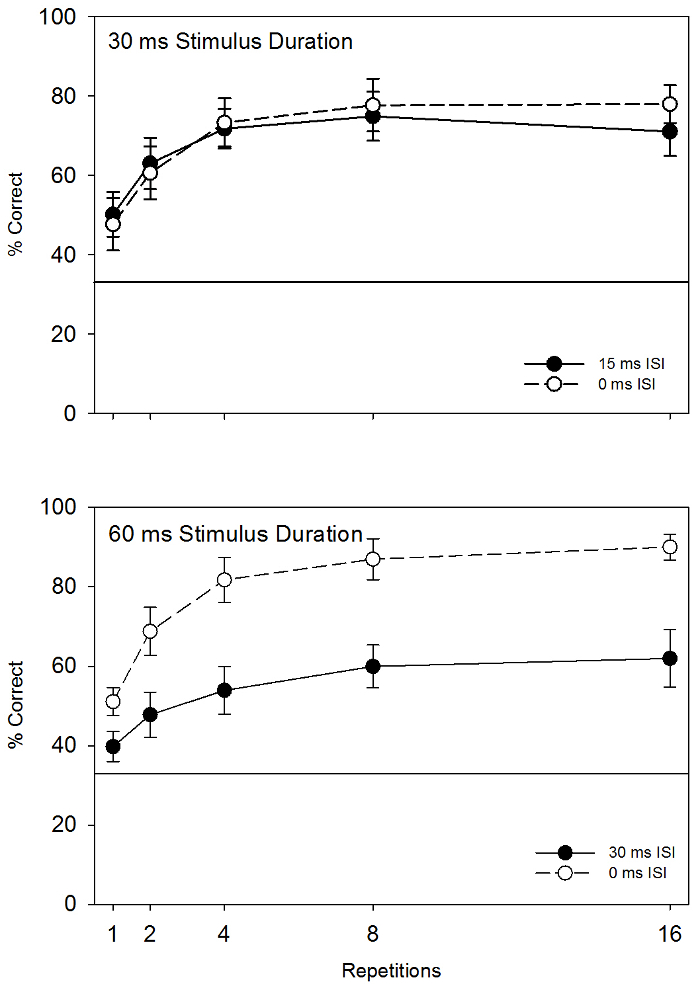
Figure 4: Rates of accurate change detection in two conditions from Experiment 2 of Herbranson and Davis, 201630. Error bars represent one standard error. The condition represented in the bottom panel produced a significant change blindness effect (better accuracy on trials without an ISI), whereas the condition represented in the top panel did not. Please click here to view a larger version of this figure.
