Healthy human fibroblasts, non-treated dysferlinopathy patient fibroblasts, and patient fibroblasts transfected with a plasmid containing the full-length dysferlin sequence were subjected to two-photon laser wounding to assess membrane resealing ability in real time. Healthy human fibroblast cells displayed low levels of FM 4-64 fluorescence activation following laser wounding and non-treated patient fibroblasts exhibited a high degree of relative fluorescence intensity following injury (Figure 5&6). Patient cells that were transfected with full-length dysferlin plasmid showed reduced relative FM 4-64 fluorescence intensity compared with non-treated patient controls and were comparable with fluorescence values observed in the healthy control (Figure 5&6).
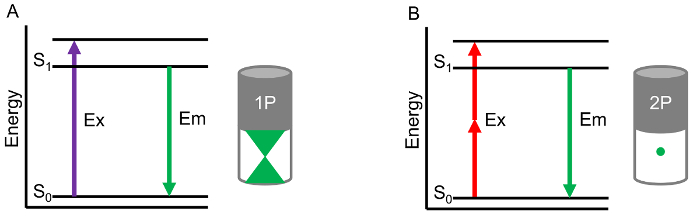
Figure 1. One-photon versus two-photon microscopy. In a traditional one-photon microscope system, a single high energy photon (UV or visible spectrum) is used to excite a fluorophore (A). In a two-photon system, two lower-energy photons (near infrared) are used to excite a fluorophore (B). Depicted within the cylinders is the fluorescence signal generated throughout the focal plane, demonstrating how the two-photon laser system can focus excitation light to within a specific region (B) and exclude light from out-of-focus planes (A). Please click here to view a larger version of this figure.
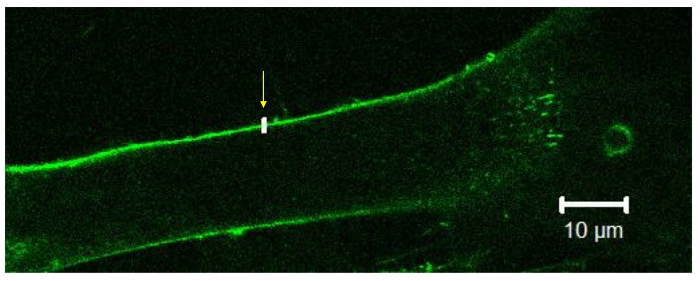
Figure 2. Placing a target at the cell membrane. Using software, a 0.2 µm x 2 µm target is drawn and placed overlapping and perpendicular to the cell membrane.The arrow indicates the target. Please click here to view a larger version of this figure.
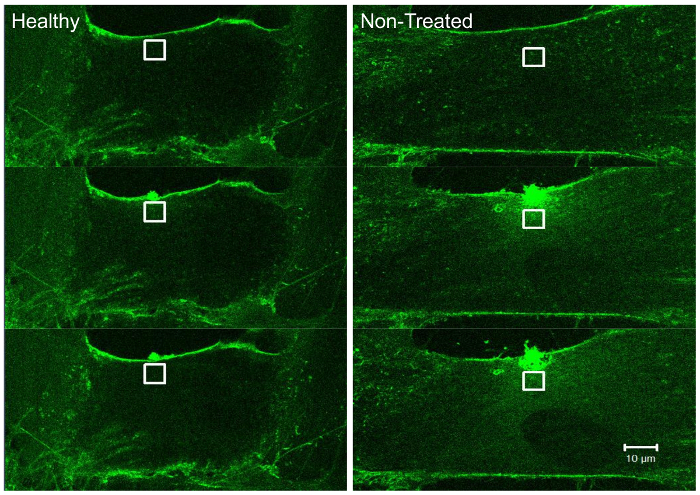
Figure 3. Fluorescence signal from FM dye as it infiltrates the cell through membrane rupture. As the cell membrane ruptures, lipophilic FM 4-64 dye infiltrates the cell and fluoresces as it binds negatively-charged phospholipids within the cytoplasm. Using software, a region of interest (ROI) can be drawn (white box) and fluorescence values within the box can be used to calculate relative fluorescence changes within the cell. Please click here to view a larger version of this figure.
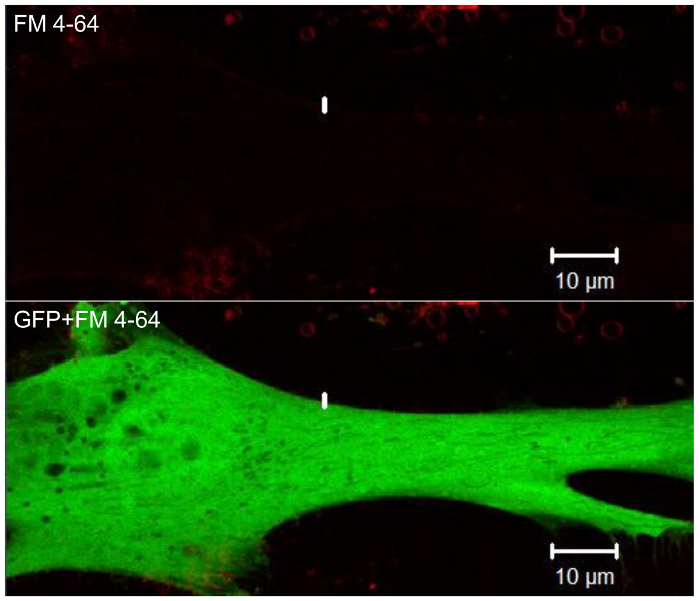
Figure 4. Fluorescence signal from GFP-conjugated full-length DYSF plasmid. Following plasmid transfection, patient fibroblast cells express full-length DYSF protein. Please click here to view a larger version of this figure.
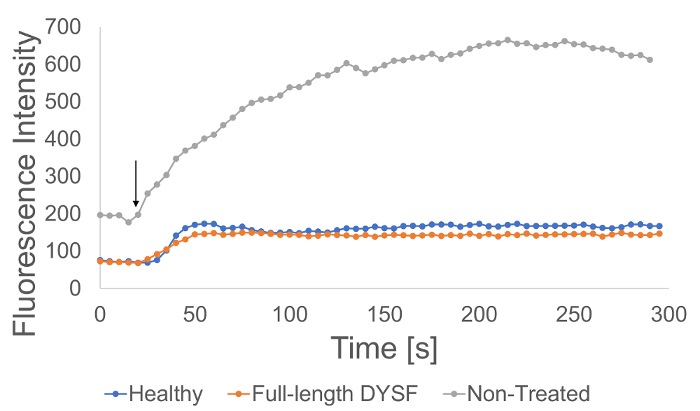
Figure 5. Quantification of fluorescence over time. Fluorescence values calculated by computer software from within the ROI can be used to determine relative fluorescence changes over time for each treatment group. The arrow indicates the time of laser ablation. Please click here to view a larger version of this figure.
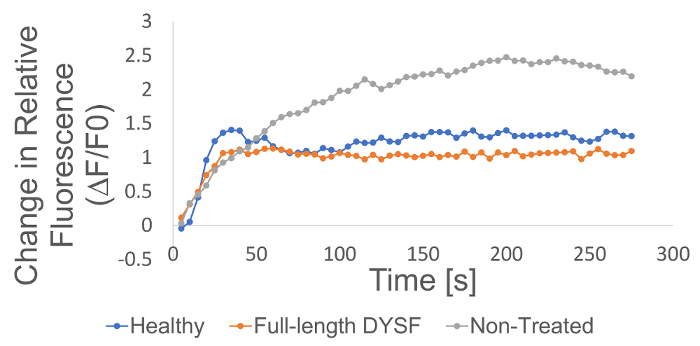
Figure 6. Change in relative fluorescence over time. Relative fluorescence (RF) values for each time point are calculated by subtracting the background fluorescence value (calculated as the mean of the fluorescence values from the timepoints prior to membrane wounding) from the given fluorescence values (Signal Intensity – Background = ΔF) and dividing the net increase (ΔF) by the fluorescence value at t = 0 (F0). Therefore, RF = ΔF/F0. Please click here to view a larger version of this figure.
