The simplified scheme of protocol for neurogenesis induction in P19 cell line is presented in Figure 1. In order to define the character of the P19 cell line in an undifferentiated state and during neurogenesis, the RT-PCR (reverse transcription-polymerase chain reaction) method was used. The undifferentiated P19 cell line expressed the pluripotency genes such as organic cation/carnitine transporter4 (Oct4) and Nanog homeobox (Nanog). Neurogenesis induced by cells aggregation in suspension culture in the presence of RA led to a rapid decrease of Oct4 and Nanog expression. In contrary, expression of neuron markers: microtubule-associated protein 2 (Map2), NeuN (also known as RNA binding protein, fox-1 homolog 3 (Rbfox3)) increased after triggered neurogenesis (Figure 2)6,14,15,22. The primers used for each gene are indicated along with nucleotide sequencesand the size of the product in Table 1. A microscopic image of the undifferentiated P19 cell line presented a round-shaped morphology (Figure 3A). After induction of neurogenesis, the neuronal structure of the cells was clearly visible 4 days after plating (Figure 3B). Additionally, Figure 4 represents the fluorescence image of MAP2 expression in the differentiated P19 cell line (4 days after plating cells)14.
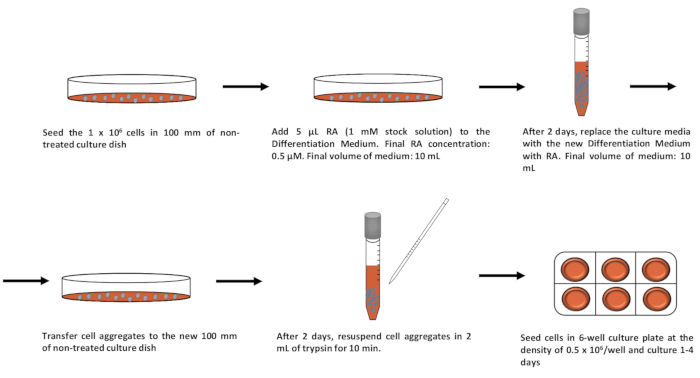
Figure 1: Protocol schematic for induction of neurogenesis in P19 embryonal carcinoma cells. Neurogenesis is induced by culturing the P19 cell line in a 100 mm non-treated culture dish with 5% of FBS and 0.5 µM RA. After 4 days, the cell aggregates are dissociated with trypsin and seeded on adherent cell culture plate for following next 4 days. Please click here to view a larger version of this figure.
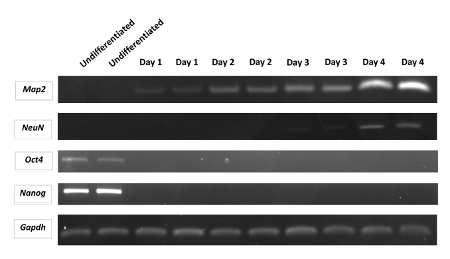
Figure 2: Changes of gene expression in P19 cell line. The band graph represents gene expression for undifferentiated P19 cell line (Oct4, Nanog) and during neurogenesis (Map2, NeuN). Glyceraldehyde-3-phosphate dehydrogenase (Gapdh) was used as the reference gene. Samples are loaded in the agarose gel (1.5%) in double replications. Abbreviations: Undifferentiated represents the undifferentiated P19 cell line without RA treatment; Day 1-4 represents subsequent days after cell plating- following 4 days after RA treatment and cell aggregation stage. Please click here to view a larger version of this figure.
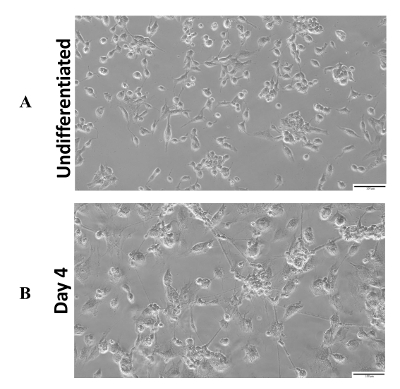
Figure 3: Representative images of analysis of P19 cell line. (A) Light microscopic images of undifferentiated P19 cell line. (B) Light microscopic images of P19 cell line after 4 days of neurogenesis- following 4 days after RA treatment and cell aggregation stage. Scale bar = 100 µm. Please click here to view a larger version of this figure.
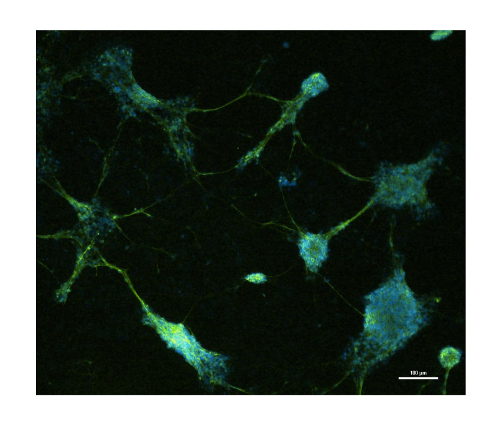
Figure 4: Representative immunofluorescence image of differentiated P19 cell line. Merged immunofluorescence image of P19 cell line stained with anti-MAP2 and DAPI at 4 days after plating. Scale bar = 100 µm. Please click here to view a larger version of this figure.
| Primer | Primer sequence | Product size (bp) |
| Gapdh | F: TGACCTCAACTACATGGTCTACA R: CTTCCCATTCTCGGCCTTG |
85 |
| Map2 | F: GCTGAGATCATCACACAGTC R: TCCTGCCAAGAGCTCATGCC |
211 |
| Oct4 | F: GGCGTTCTCTTTGGAAAGGTGTTC R: CTCGAACCACATCCTTCTCT |
313 |
| NeuN | F: GGCAAATGTTCGGGCAATTCG R: TCAATTTTCCGTCCCTCTACGAT |
160 |
| Nanog | F: AAAGGATGAAGTGCAAGCGGTGG R: CTGGCTTTGCCCTGACTTTAAGC |
520 |
Table 1: Primers used for RT-PCR.
