Figure 1 and Figure 2 show representative results of immunofluorescence localization of DNM in the mouse caput epididymis. Each of the three DNM isoforms investigated display distinct localization profiles. Thus, DNM1 is characterized by relatively modest diffuse labeling of the epididymal cells throughout the initial segment and caput epididymis (Figure 2A). By contrast, the DNM2 isoform was first detected in the vicinity of the opposing basal and apical border of cells in the initial segment, before being repositioned to the supranuclear domain in cells within the adjacent downstream caput segment (i.e., zones 2 – 5) (Figure 1B, C). Notably, however, the intensity of DNM2 labeling gradually decreased between zones 2 to 5 of the caput epididymis, a result that essentially mirrors the secretory activity of these epididymal segments21 (Figure 1B, C). Accordingly, the supranuclear labeling of DNM2 was subsequently shown to correspond to the distribution of the Golgi apparatus within caput principal cells21. Spermatozoa isolated from the same epididymal region showed intense acrosomal labeling for DNM2 (Figure 1D). As a caveat, however, equivalent DNM2 labeling was not routinely detected on luminal spermatozoa within our tissue sections. This phenomenon is the one we have encountered on several occasions when applying a range of antibodies targeting different epididymal/sperm antigens and presumably arises due to issues associated with antigen presentation and/or masking in the paraffin-embedded tissue sections. In any case, such differences emphasize the importance of conducting parallel immunofluorescent labeling of isolated spermatozoa alongside that of the epididymal tissue itself. Differing from both DNM1 and DNM2, the DNM3 isoform was mainly detected in the apical domain of a small number of caput epithelial cells (Figure 2B, green arrows), which were shown to correspond to the clear cell sub-population by co-labeling with the recognized clear cell marker, ATP6V1B1 (Figure 2B, red arrows). In a similar manner, representative markers that have proven suitable for differentiating the different epididymal epithelial cell types are summarized in Table 229,30,31,32,33,34.
In addition to the description of the techniques for the subcellular localization of proteins residing within the epididymal epithelium, here, we also report our recently optimized protocols for the study of secretory proteins encapsulated within epididymosomes, small extracellular vesicles that represent an important component of the luminal milieu responsible for supporting sperm maturation and storage22. Combined, step 2 and Figure 3 provide a detailed step by step account of the methodology used for the isolation of highly enriched populations of epididymosomes from mouse caput epididymal tissue. Notably, however, these methods are readily applicable for the isolation of alternate populations of epididymosomes originating from more distal epididymal segments. Owing to the potential for contamination of these samples, we also described the stringent characterization protocols that we routinely employ for each epididymosome preparation. These include the assessment of the size and heterogeneity of the epididymosome populations using both high-resolution electron microscopy and dynamic light scattering techniques. In tandem, we also utilize immunoblotting strategies to assess the enrichment of recognized extracellular vesicle markers and the corresponding absence of proteins that are characteristic of potential contaminants (i.e., anti-hemoglobin (HBB) as a marker of blood contamination and anti-arachidonate 15-lipoxygenase (ALOX15) and anti-IZUMO1 antibodies as markers of cytoplasmic droplet and sperm contamination, respectively)22. Although we have found that the contaminants are rare, if they are encountered, we immediately discard the epididymosome preparation.
The non-overlapping localization of DNM isoforms in the caput epididymis prompted a further investigation of their potential roles in regulating the epididymal microenvironment. For this purpose, an immortalized mECap18 cell line was utilized as an in vitro model to study epididymal cell secretory activity. Previous characterization of this cell line has shown that it harbors a mixed cell population, which stain positive for either principal or clear cell markers. Moreover, mECap18 cells have also proven suitable for reporting physiological profiles of epididymal gene and protein expression under different in vitro treatment regimens35. Prior to use, DNM localization was assessed in cultured mECap18 cells by settling these onto poly-L-lysine treated coverslips (Figure 5A) and subjecting them to immunofluorescence detection. Consistent with the distribution patterns recorded in caput epididymal tissue sections, DNM1 was detected throughout the cytoplasm of mECap18 cells, while DNM2 was concentrated within the supranuclear domain of these cells and DNM3 was characterized by discrete foci of membrane staining within a small sub-population number (i.e., 11%) of the mECap18 cells which were ATP6V1B1 positive (Figure 5B). These data affirm the utility of the mECap18 cell line as a valuable resource for investigating the role of DNM in regulating epididymal cell secretory/absorptive activity.
Accordingly, step 4 describes the methodology for the analysis of mECap18 cell secretory activity; the techniques which are broadly amenable for assessing the impact of a range of different experimental conditions. In our study, we applied selective pharmacological interventions to suppress the activity of DNM1 and DNM2 prior to the visualization and quantification of the profile of proteins released from mECap18 cells into conditioned medium21. An important feature of this analysis, however, was to ensure that mECap18 cells were thoroughly washed and cultured in the absence of FBS supplementation. Whilst such a step was essential to preclude the contamination of conditioned medium with FBS derived proteins, it nevertheless carries the attendant risk of negatively impacting mECap18 cell growth and/or viability. In controlling for this possibility, we noted that the mECap18 cell line tolerated FBS free culture and the introduction of DNM inhibitors for the duration of our incubation window (i.e., 12 h). Indeed, over this time course, cell viability remained above 90% in all experimental replicates. This approach could therefore serve as a useful proof-of-concept strategy to identify the function of specific epididymal proteins before committing to investment into gene manipulation strategies.
| Heat induced epitope retrieval solution | 10 mmol/L sodium citrate | 50 mmol/L Tris (pH 10.5) |
| Time | 3 min | 3 min |
| 6 min | 6 min | |
| 9 min | 9 min | |
| 12 min | 12 min |
Table 1: General conditions for the optimization of heat-induced antigen retrieval for the use with paraffin-embedded epididymal sections. The fixation process can be problematic as different epitopes often require the use of different fixation techniques, thereby necessitating that the methodology is optimized for each antigen.
| Epithelial cell type | Distribution | Marker | References (PMID) |
| Principal cell | Whole epididymis | AQP9 | 11027599, 17360690 |
| Clear cell | Caput, corpus and cauda | V-ATPases, CIC-5 | 19448084, 12475763 |
| Basal cell | Whole epididymis | CLDN1 | 11159859, 21441423 |
| Narrow cell | Initial segment | V-ATPases, CIC-5 | 19448084, 12475763 |
Table 2: Representative markers suitable for the detection of different primary epididymal epithelial cell types.
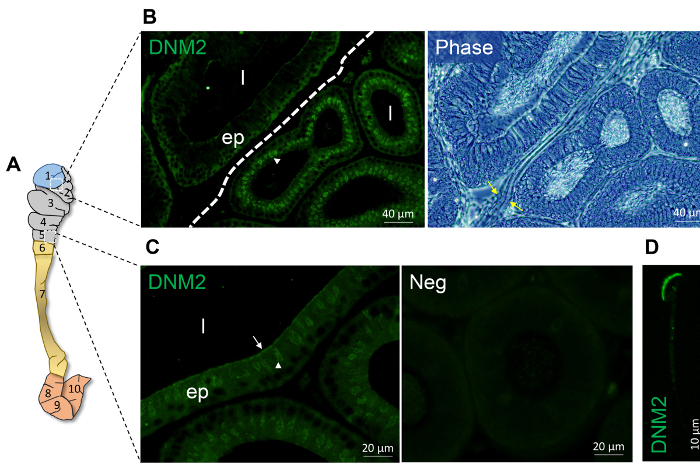
Figure 1: Spatial expression of DNM 2 within the proximal mouse epididymis. (A) Schematic model of epididymis depicting the partitioning on the mouse epididymis into 10 zones physically separated septa as reported by Turner and collegues20. In this model, zone 1 corresponds to the initial segment, zones 2-5 correspond to the caput epididymis, zones 6-7 correspond to the corpus epididymis and zones 8-10 represent the cauda epididymis. (B-C) Immunofluorescence localization of DNM2 revealed zone-specific distribution patterns (indicated by white arrowhead and arrow). The border between zone 1 and 2 is demarcated by a dotted line or denoted by yellow arrows. (D) DNM2 is also expressed in the peri-acrosomal domain of spermatozoa isolated from the caput epididymis. However, no such staining was routinely detected in luminal spermatozoa within the corresponding epididymal sections. ep, epithelial cells; l, lumen; Neg, secondary antibody only control. Experiments were replicated on material from three animals and representative immunofluorescence images are presented. Please click here to view a larger version of this figure.
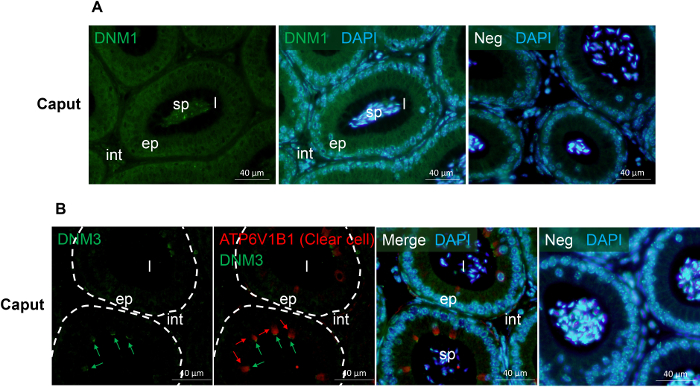
Figure 2: Immunofluorescence detection of DNM 1 and DNM 3 in the mouse caput epididymis. (A) The localization of DNM136 was examined in the mouse caput epididymis. (B) Co-localization of DNM336 and the clear cell marker, ATP6V1B137 in the mouse caput epididymis. This analysis confirmed that both DNM3 (green arrows) and ATP6V1B1 (red arrows) reside in the clear cell sub-population but display minimal sub-cellular overlap. ep, epithelial cells; int, interstitium; l, lumen; sp, sperm; Neg, secondary antibody only control. Cell nuclei were counterstained with DAPI (blue). Experiments were replicated on material from three animals and representative immunofluorescence images are presented. Please click here to view a larger version of this figure.
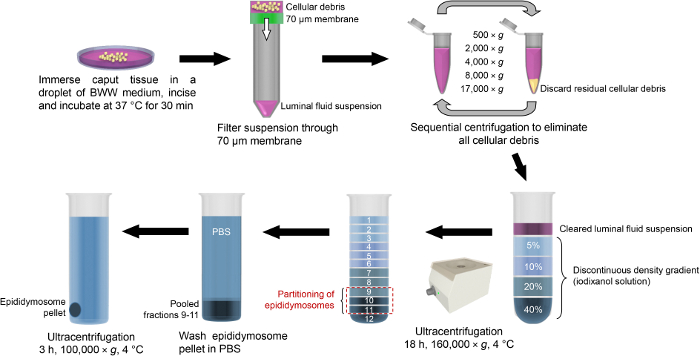
Figure 3: Schematic of isolation protocols used for enrichment of mouse caput epididymosomes. After the dissection, caput epididymal tissue is immersed into a droplet of BWW medium and incised to release the luminal contents. The luminal fluid is then filtered through a 70 µm membrane and the resultant suspension is centrifuged at increasing velocity in order to pellet any residual cell debris. The cleared suspension is then loaded atop of a discontinuous density gradient (iodixanol solution) and subjected to overnight ultracentrifugation. Epididymosomes partition into fractions 9 – 11, which are pooled, washed via dilution into PBS and returned to the ultracentrifuge to pellet the epididymosomes. Please click here to view a larger version of this figure.
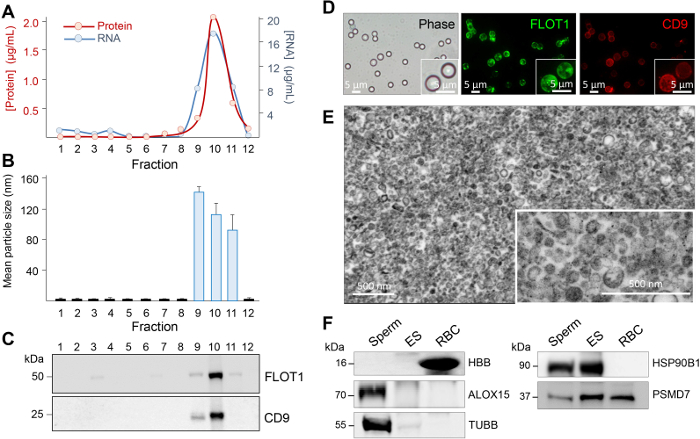
Figure 4: Assessment of epididymosome purity. Twelve equal fractions were recovered after the ultracentrifugation of the gradient and an aliquot of each prepared for (A) protein and RNA quantification, (B) size heterogeneity assessment by using dynamic light scattering, and (C) immunoblot analysis of epididymosome marker distribution. Additional characterization steps included (D) dual-labeling of epididymosomes concentrated onto aldehyde/sulphate latex beads, (E) transmission electron microscopy assessment, and (F) immunoblot assessment of spermatozoa (Sperm) and red blood cell (RBC) contamination by using either anti-arachidonate 15-lipoxygenase (ALOX15, cytoplasmic droplet/sperm contamination) or anti-hemoglobin (HBB, RBC contamination). Immunoblots were also probed with known epididymosome cargo (26S proteasome non-ATPase regulatory subunit 7, PSMD7; heat shock protein 90kDa beta member 1, HSP90B1; and beta tubulin, TUBB). These data were originally published in Scientific Reports (PMID: 27549865) and have been reproduced here with the permission of the publisher, Springer Nature. Please click here to view a larger version of this figure.
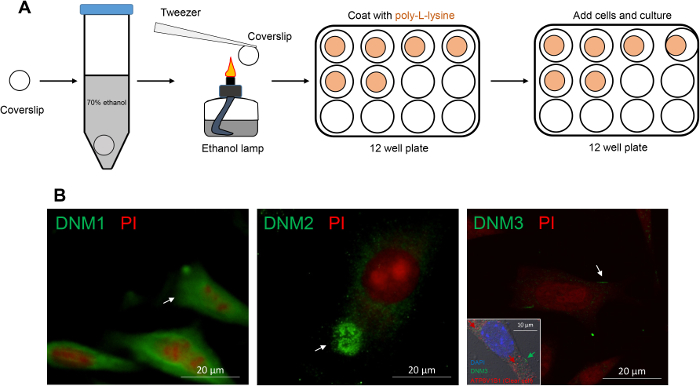
Figure 5: Immunofluorescence detection of DNM isoforms in mECap18 cells reveal distribution patterns that accord with those detected in caput epididymal tissue. (A) Schematic of coverslip preparation for sterile mECap18 cell culture. (B) Representative immunofluorescence images of DNM staining revealed cellular distribution patterns (arrows and inset (dual labeling of DNM3 and clear cell marker ATP6V1B1)) that mirrored those detected within epididymal tissue sections. Cell nuclei were counterstained with either propidium iodide (PI; red) or DAPI (blue). Experiments were replicated on material from three animals and representative immunofluorescence images are presented. Please click here to view a larger version of this figure.
