Using the described techniques (summarized in Figure 1), retinal explants from 8 week-old wild type (WT) mice were compared to age- and background-matched transducin null mice (Gnat1-/-). Because Gnat1-/- animals lack the machinery to close cyclic-nucleotide gated ion channels in response to light stimuli, their rod photoreceptors remain depolarized even in light14. The subsequent need to maintain potassium efflux would create a large ATP demand, resulting in bioenergetic strain. To determine if such shifts in energy demands would increase oxidative phosphorylation or glycolytic flux, tissues from wild type mice and Gnat1-/- mice were compared using the extracellular flux analyzer. At baseline, in the presence of 5 mM glucose and 1 mM pyruvate, retinal tissues from wild-type animals have similar rates of acid efflux compared to Gnat1-/- mutants. Similar patterns are seen after addition of 20 mM glucose and the glycolytic inhibitor 2-DG (Figure 2A). These data may also be mathematically transformed to represent fractional changes from baseline (Figure 2B), a format which may allow for better comparison between different experimental interventions.
Absolute OCR at baseline is equivalent between WT and mutant retinal tissue (Figure 3A). The addition of 20 mM glucose increases mitochondrial respiration, but no change is observed between groups in terms of absolute quantification or in terms of change from baseline (Figure 3B). The addition of 1mM FCCP (an uncoupling agent) to demonstrate maximal mitochondrial respiratory rates does not significantly increase OCR above the level seen with high glucose in WT or Gnat1-/- tissues. However, signals from both mice drastically drop after the addition of RAA cocktail.
Ideally, extracellular flux experiments run in the presence of the ATP synthase inhibitor oligomycin aid in identifying sources of proton leak, occurring when movement through the electron transport chain is not associated with ATP production16. In retinal explants from 8 week-old C57BL/6J mice, oligomycin treatment robustly lowers OCR, as expected (Figure 4). But subsequent addition of FCCP, to find maximal OCR, only nominally increases the OCR to about 60% of baseline, consistent with a prior study11.
The XF24 analyzer uses submersible probes that make a tight seal within the tissue well, creating a transient microenvironment for measurement of oxygen and acid flux. Positioning of retinal tissue in the well plate (in relation to the sensors) could potentially influence OCR recordings, and lead to unwanted confounders, and some reserachers have advocated placing retinal tissue directly underneath the oxygen sensor with the photoreceptor outer segments oriented toward the sensor11. To test the effects of retinal tissue position on OCR measurements, tissues from young C57BL/6J mice (8 weeks old) were analyzed in two orientations: retinal ganglion cells facing down onto the plate (i.e., photoreceptors placed upright closest to sensor) or facing up toward the sensor. No differences in relative OCR in response to FCCP or RAA were observed, indicating equivalent sensitivity for maximal and minimal mitochondrial respiratory rates in either orientation (Figure 5A). In addition, absolute OCR measurements were also equivalent between retinal tissues in either orientation (Figure 5B).
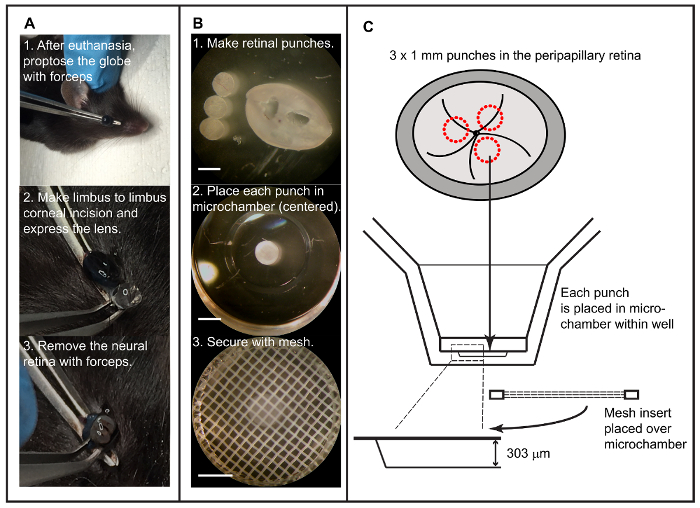
Figure 1: Isolation of retinal explant and setup into the extracellular flux analyzer. A. Isolation of the neural retina in situ by applying propulsive force on globe with forceps, incising the cornea, and removing retina after discarding lens and anterior hyaloid membranes. B. Preparation of tissue for flux recording with retinal punch creation, placement of individual punches into islet capture microplate, and use of a mesh insert to minimize tissue movement with microchambers (scale bars = 1 mm). C. A schematic, derived from manufacturer-provided data, showing the procedure outline and demonstrating that height of the microchamber is adequate to accommodate the murine retina. Please click here to view a larger version of this figure.
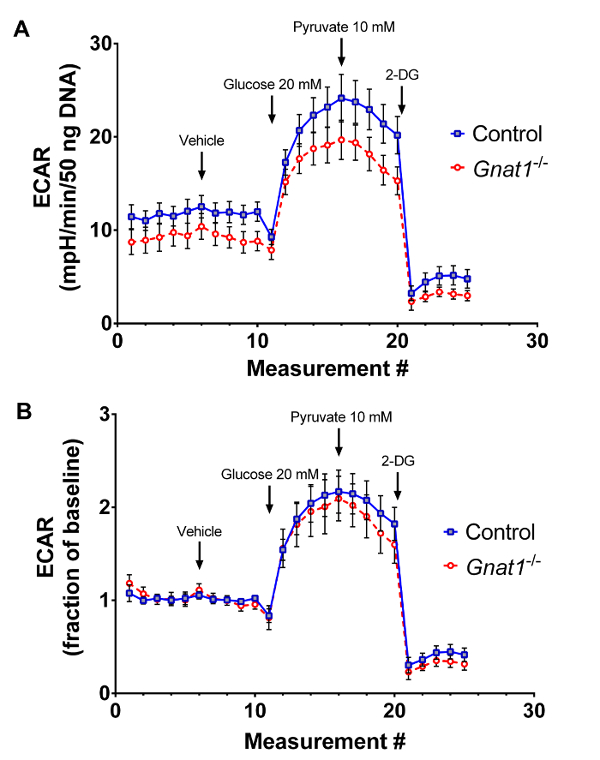
Figure 2: Mitochondrial stress analysis using explanted retina from 8 week-old animals. A. Averaged tracings with standard error of measurement (SEM), normalized to DNA content of the tissue, from an experiment optimized for oxygen consumption recordings, comparing transducin knockout animals (Gnat1-/-) to wild type controls. B. Same experiment with data transformed such that baseline recordings are set as the reference. Please click here to view a larger version of this figure.
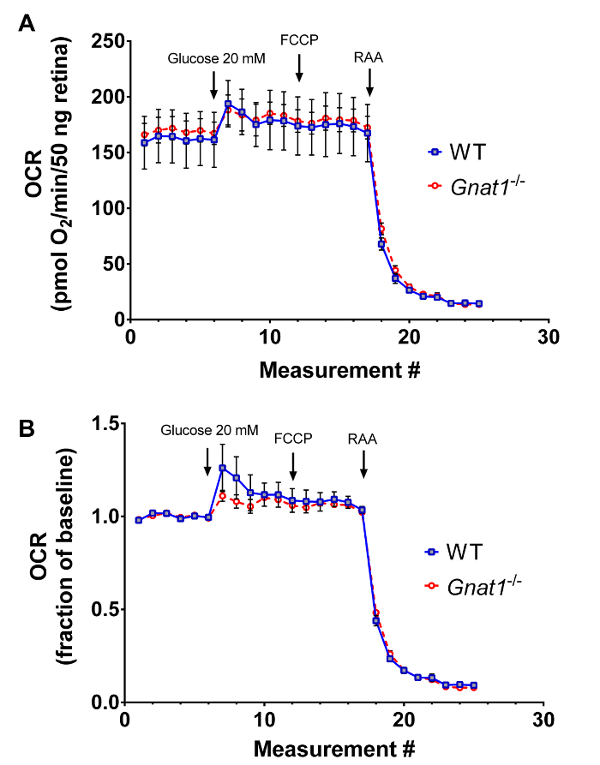
Figure 3: Analysis of glycolytic rate using explanted retina from 8-week old animals. A. Averaged, DNA content-normalized, tracings from an experiment optimized for acid efflux recordings comparing transducin knockout animals (Gnat1-/-) to wild type controls. B. Data from the same experiment normalized to baseline recordings. Traces show mean ± SEM. Please click here to view a larger version of this figure.
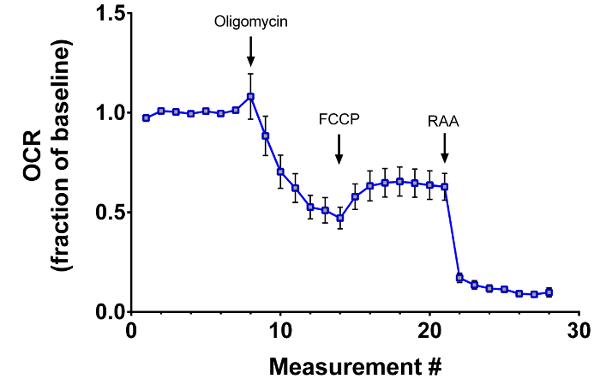
Figure 4: Irreversible effects of oligomycin on retinal respiratory rates. In acutely prepared retinal explants from 8 week-old C57BL/6J mice, oligomycin treatment reduces the maximal OCR induced by subsequent addition of FCCP. Traces show mean ± SEM. Please click here to view a larger version of this figure.
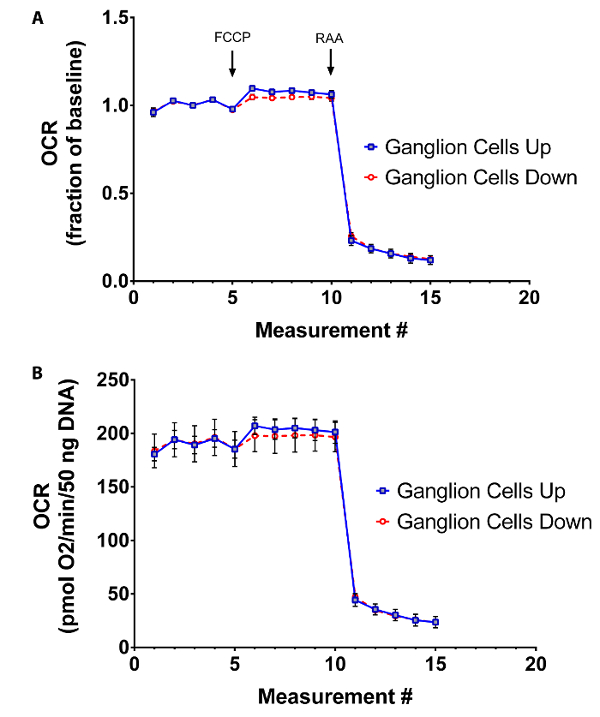
Figure 5: Effects of tissue positioning on OCR measurement. Retinal tissues from 8 week-old C57BL/6J mice were placed in wells with ganglion cells facing toward the sensor (Ganglion Cells Up) or away from the sensor (Ganglion Cells Down). Between groups, similar extracellular flux recordings are observed in terms of (A) respiratory capacity relative to baseline or (B) in terms of absolute quantification. Traces show mean ± SEM. Please click here to view a larger version of this figure.
