Members of the mitis group S. mitis, S. oralis, and S. gordonii rapidly killed the worms, as opposed to S. mutans, S. salivarius, and non-pathogenic E. coli OP50 (Figure 3A). The median survival for S. mitis, S. oralis, and S. gordonii was 300 min, 300 min, and 345 min, respectively. To determine if the killing was mediated by H2O2, catalase was supplemented to THY agar. The killing of the worms was abolished in the presence of catalase (Figure 3B). To further confirm whether streptococcal derived H2O2 mediated killing of the worms, survival on the ΔspxB mutant strain, WT strain, and complement strain ΔspxB;spxB+ of S. gordonii was analyzed. Death of the worms was not observed on the ΔspxB mutant strain compared to the wild-type and complement strains (Figure 3C). These data suggest that the H2O2 produced by the mitis group mediates killing of the worms. We also observed similar killing kinetics when the worms were exposed to clinical isolates of the mitis group streptococci obtained from the blood of cancer patients (Figure 3D). Based on the data, the pathogenicity caused by H2O2 produced by the mitis group streptococci was assessed.
To identify host genes that are essential against streptococcal infections, skn-1 was knocked down, which encodes for the oxidative stress response transcription factor in C. elegans. Then, survival relative to the vector control treated worms was compared. A significant decrease in the survival of the skn-1 knockdown worms was observed compared to the vector control treated worms (Figure 4A). This data was further validated using a skn-1 mutant strain, and its survival was compared to that of the N2 wild-type worms. We observed a similar killing phenotype as the skn-1 mutant, as seen with the skn-1 knockdown, demonstrating that SKN-1 influenced the survival of the worms on the mitis group (Figure 4B).
Next, it was determined whether the H2O2 produced by the mitis group caused localization of SKN-1B/C::GFP in the worms. Localization of SKN-1B/C::GFP was observed in worms exposed to the wild-type and complement stains and not in response to the ΔspxB mutant strain of S. gordonii (Figure 5A,B). Furthermore, to determine the activation of SKN-1, components of the p38 MAPK pathway were knocked down. Reduced localization of SKN-1B/C::GFP in nsy-1, sek-1, pmk-1, and skn-1 knockdown worms relative to the vector control treated worms was observed. The data suggests the p38 MAPK is required for the activation of SKN-1 in response to H2O2 produced by the mitis group (Figure 5C,D).
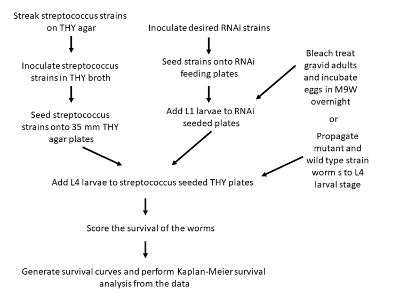
Figure 1: Flowchart depicting the steps involved in preparation of the survival assays. Please click here to view a larger version of this figure.
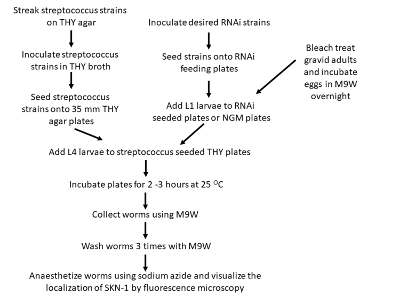
Figure 2: Flowchart depicting the steps involved in localization of SKN-1 during infection. Please click here to view a larger version of this figure.
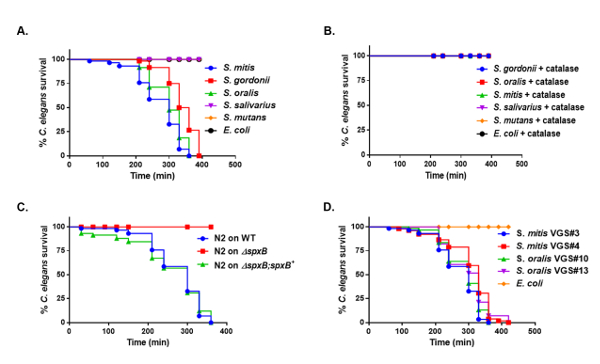
Figure 3: H2O2-mediated killing of C. elegans by mitis group streptococci. Kaplan-Meier survival curves of L4 larvae exposed to (A) S. gordonii, S. oralis, S. mitis, S. salivarius, S. mutans, and E. coli OP50. (B) S. gordonii, S. oralis, S. mitis, S. salivarius, S. mutans, and E. coli OP50 on THY plates in the presence of 1,000 U of catalase. (C) S. gordonii WT, ΔspxB mutant, and ΔspxB;spxB+ complement strains on N2 L4 larvae. (D) S. oralis (VGS#3), S. oralis (VGS#4), S. mitis (VGS#10), S. mitis (VGS#13), and E. coli OP50. The data are representative of experiments repeated two or more times, with n = 60 worms for each condition. Kaplan-Meier log rank analysis was used to compare survival curves and calculate the median survival. P values < 0.05 were considered to be statistically significant. This figure has been modified and adapted with permission15. Please click here to view a larger version of this figure.
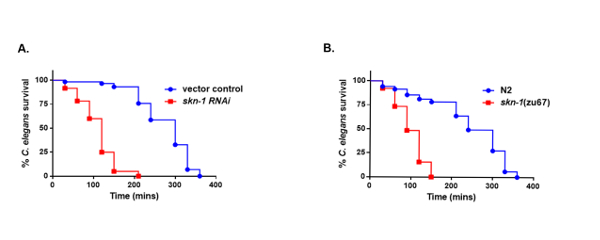
Figure 4: SKN-1 is required for survival of the worms on S. gordonii. (A) Survival of vector control treated and skn-1 knockdown worms exposed to S. gordonii. (B) Survival of N2 and skn-1(zu67) mutant worms fed on S. gordonii. The data are representative of experiments repeated two or more times, with n = 60 worms for each condition. Kaplan-Meier log rank analysis was used to compare survival curves and calculate the median survival. P values < 0.05 were considered to be statistically significant. This figure has been modified and adapted with permission15. Please click here to view a larger version of this figure.
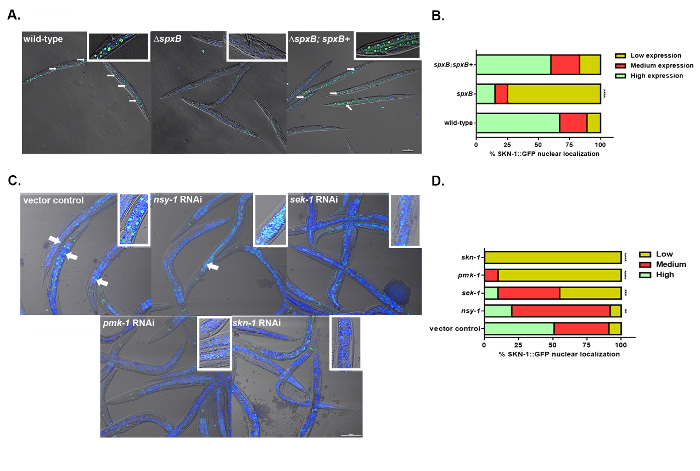
Figure 5: Streptococcal H2O2 mediated activation of SKN-1 is dependent on the p38 MAPK pathway. (A) Representative images of the localization of SKN-1B/C::GFP in worms exposed to the WT, ΔspxB mutant, and ΔspxB;spxB+ complement strains of S. gordonii. Closeups are shown in the upper righthand corners of each image. Scale bar = 100 µm. (B) The degree of nuclear localization of SKN-1B/C::GFP and percentage of worms in each category fed on WT, ΔspxB mutant, and ΔspxB;spxB+ complement strains of S. gordonii. Significantly low levels of nuclear localization of SKN-1B/C::GFP were observed in the ΔspxB mutant (p < 0.0001) compared to the WT and ΔspxB;spxB+ complement strains of S. gordonii. (C) Representative images of the localization of SKN-1B/C::GFP in nsy-1, sek-1, pmk-1, skn-1 knockdown, and vector control treated worms on S. gordonii. Closeups are shown in the upper righthand corners of each image. Scale bar = 100 µm. (D) The degree of SKN-1B/C::GFP nuclear localization and percentage of worms in each category fed on nsy-1, sek-1, pmk-1, skn-1 knockdown, and vector control treated worms on S. gordonii. Significantly low levels of nuclear localization of SKN-1B/C::GFP were observed in the nsy-1 (p < 0.01), sek-1 (p < 0.001), pmk-1 (p < 0.0001),and skn-1 knockdown (p < 0.0001) compared to the vector control treated worms on S. gordonii. Greater than 100 worms exposed to each strain were imaged, and the experiment was repeated three times. This figure has been modified and adapted with permission15. Please click here to view a larger version of this figure.
