The representative results presented in this paper were derived from 15N-tracing experiments of salt marsh sediments. The sampled salt marsh was newly created in the aftermath of the 2011 Great East Japan Earthquake in the Moune area of Kesen-numa city in Miyagi Prefecture, Japan. In September 2017, surface sediments (0–3 cm) were collected at two sites in the subtidal and intertidal zones. First, immediately after collection, the sediment was sieved through a 4-mm mesh to remove plant roots, shells debris, and rubble and then homogenized. The samples were stored at 4 °C until the DNRA analysis was conducted.
The incubation procedures for the 15NO3− and the simultaneous determination of the 14NH4+ and 15NH4+ concentrations were carried out as described in the protocol section. An increase in the 15NH4+ concentration throughout the incubation period was observed for all sediments (Figure 2). We calculated the DNRA rates by dividing the accumulation rate of 15NH4+ by the isotope ratio of the NO3− pool29. The calculated rates were within the range of 24.8–177 nmol-N g−1 dry soil h−1 (Table 3) and were comparable to values found in previous studies. This range of obtained rates is higher than the reported values derived from similar environments including those from intertidal sediments17, salt marshes5,16, and other estuarine environments18,33,34, as well as from eutrophic environments such as a shallow river estuary in North Carolina31 and the Shanghai urban river networks32. Conversely, Fernandes et al.13 reported higher potential DNRA rates in organically rich mangrove soils in India. In general, DNRA is thought to be favored by a high ratio of available C to electron acceptors35,36,37. The samples demonstrating the representative results were taken from a salt marsh newly created by an earthquake, which had originally been used as a cultivation field. This particular characteristic of the samples may contribute to the observed high DNRA rate. Consistent with this speculation, the DNRA rate in the intertidal zone, which is rich in organic compounds (data not shown) compared to the subtidal zone, was higher than that in the subtidal zone (Figure 2, Table 3).
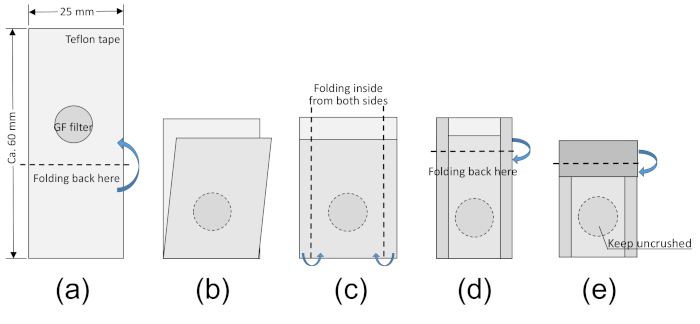
Figure 1: Preparation of a PTFE envelope for capturing gaseous NH3. The PTFE envelope used in the diffusion procedure is prepared by folding PTFE tape following the instructions shown in panels (A)–(E). The acidified filter inside the envelope captures the gaseous NH3. These steps should be conducted quickly. Detailed information is given in steps 1.2–1.7 in the protocol section. Please click here to view a larger version of this figure.
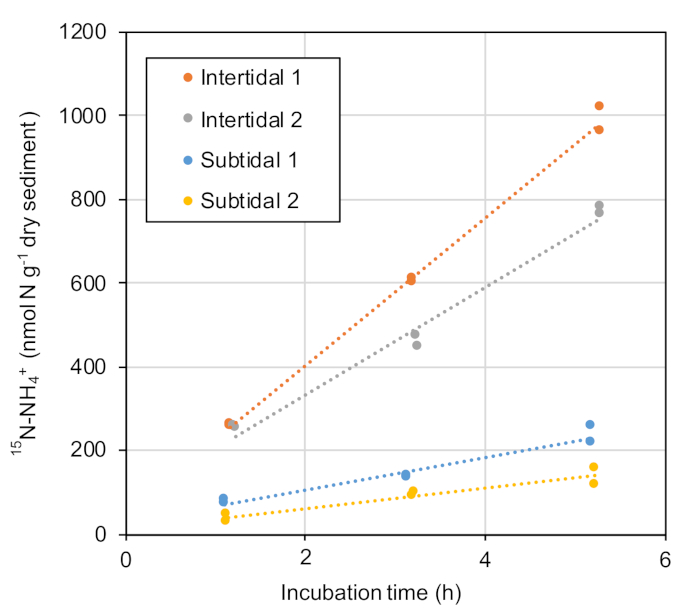
Figure 2: Change in the 15NH4+ concentration via the anaerobic incubation of sediments. The 15N tracer incubations of the sediment samples were conducted in duplicate. The concentration of 15N-NH4+ is shown in nmol per dry weight of sediment. Please click here to view a larger version of this figure.
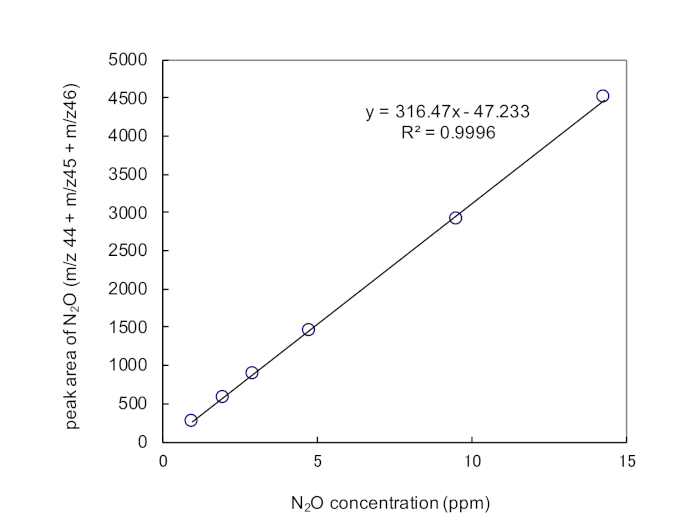
Figure 3: An example of calibration curve of low concentration N2O. Peak area of N2O was obtained by sum of the peak area of m/z 44, m/z 45, and m/z 46. Configurations for GC/MS analysis is shown in Table 2. Please click here to view a larger version of this figure.
| 15N-labeled and non-labeled substrates added to each vial | ||
| 100mM | 100mM | |
| NH4Cl | K15NO3 | |
| Volume (µL) of 100 mM stock solution added to each vial | 24 | 60 |
| Final concentration (µmol/L)* | >230§ | 570 |
| *shown values are calculated by assuming that water content of the sediment is 50% | ||
| §depending on the background ammonium concentration | ||
Table 1: Combinations of substrates amended to vials retaining approximately 11 mL of the sediment suspension. Samples were prepared in duplicate and subjected to further analyses after 1 h, 3 h, and 5 h of incubation.
| Equipment | |
| quadrupole GCMS | Shimadzu GCMS-QP2010 Ultra |
| column | CP-PoraBONDQ 25m; φ 0.32mm; film thickness, 5µm |
| Analytical conditions | |
| column temp | 40 °C |
| injection port temp | 100 °C |
| carrier gas stream | Total flow rate, 47.1 mL•min-1 flow rate in column, 2.10 mL•min-1 |
| sprit ratio | 20 |
| detection voltage | 1.5 kv |
| Sensitivity of N2O | |
| lower limit of detection (LOD)* | 1.42 pmol |
| lower limit of quantification (LOQ)* | 4.58 pmol |
| *LOD and LOQ were determined by a linear relationship among a serial dilution of N2O (0.97, 1.94, 2.91, 4.75, 9.50, 14.3 ppm) in He, corresponding responses in peak area, and S/N ratio. LOD and LOQ were calculated as concentrations equivalent to S/N=3 and S/N=10, respectively. |
Table 2: Conditions for the GC/MS analysis.
| sediment | DNRA rate | enrichment of NO3– * |
| nmol-N g-1 hr-1 | atom% | |
| Intertidal 1 | 177 | 99.9 |
| Intertidal 2 | 129 | 99.0 |
| Subtidal 1 | 39.3 | 99.9 |
| Subtidal 2 | 24.8 | 99.0 |
| *same as atom% of added KNO3; complete elimination and no nitrification under used incubation conditions was tested previously. |
Table 3: Potential DNRA rates of the tested intertidal and subtidal sediments.
