Multiple photographs showing different views of the MPP flow chamber on the microscope stage (upper panel) and a schematic representation of the MPP flow chamber (bottom panel) are shown in Figure 1. The schematic details the dimensions of the entire device and flow chamber. Figure 2 shows a photograph of the gravity perfusion system to the MPP flow chamber in our laboratory (upper panel). Also shown is a schematic representation of the flow system (bottom panel) intended to highlight the steps which isolate the cells in the flow chamber from the rush of solution from the perfusion system and from the force of the vacuum removal of solution.
A hallmark of mechanosensitive ion channels is an abrupt return to baseline levels upon cessation of mechanical stimulus3,6,7. Figure 3 shows as an example that removing the shear stress stimulus during electrophysiological recordings of Kir current from a freshly isolated endothelial cell results in a return to baseline currents initially recorded in a static bath. The return to baseline current levels after stopping flow to the MPP chamber occurred within ten seconds of flow cessation.
Whole-cell (WC) electrophysiological recordings, especially those taken from freshly isolated cells, often have obvious leak background currents that can mask channel activity. Some ion channels, such as those of the inwardly rectifying K+ channel family, have biophysical properties that allow for subtracting the leak background current for more accurate analysis. Figure 4 shows as an example the process from raw data (Figure 4A) to calculated and plotted linear outward leak (Figure 4B) to final, leak subtracted representative trace (Figure 4C). See a detailed explanation for calculating and subtracting leak from the raw perforated WC patch recording in the accompanying legend to Figure 4.
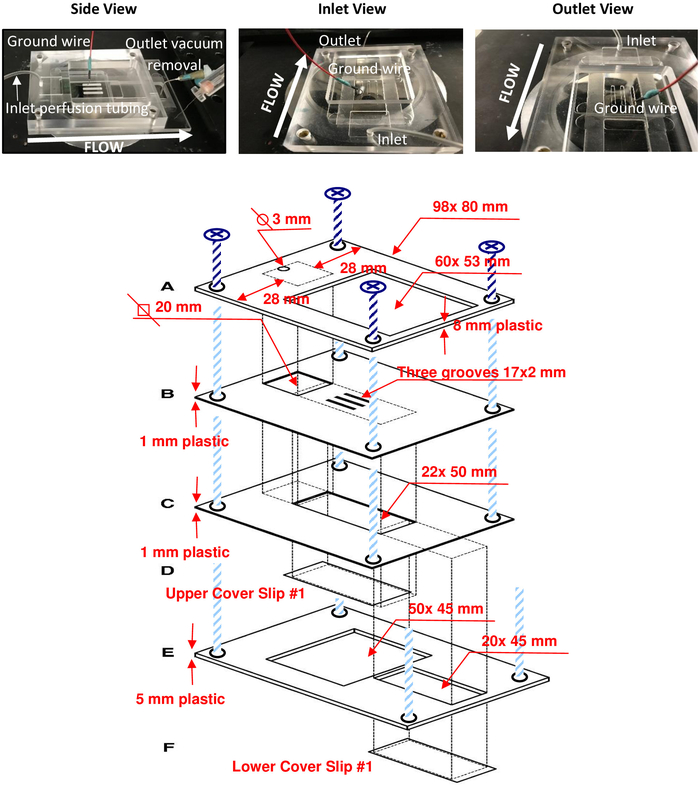
Figure 1: MPP flow chamber and detailed schematic. Photographs of the assembled MPP flow chamber (upper panel) show the chamber on the microscope stage from three different views: the side as viewed during experiments (left), from the perfusion inlet (middle), and from the vacuum outlet (right) which is out of view in the photograph. The direction of flow is labeled in each. Notice that the ground wire can easily fit into one of the 2 mm slits when bent at a 90° angle. A detailed schematic (bottom) shows exact dimensions for replication of the apparatus. Please click here to view a larger version of this figure.
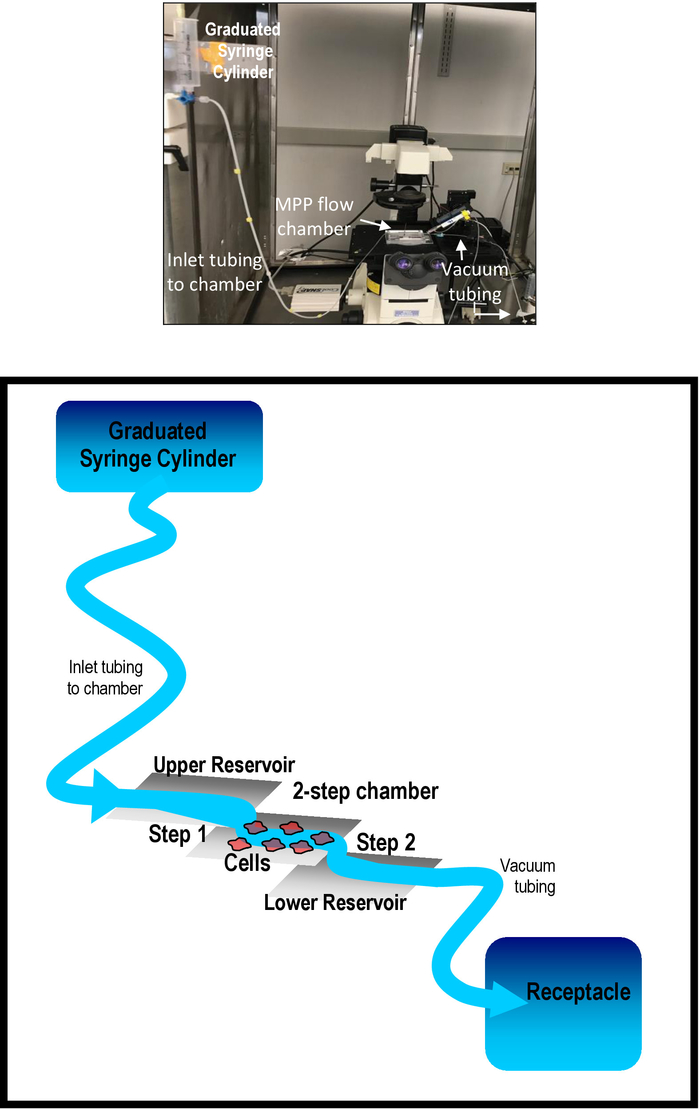
Figure 2: The gravity perfusion system. A labeled photograph of the gravity perfusion system in our laboratory is shown in the upper panel. The two-step MPP flow chamber and gravity perfusion system are detailed in the bottom panel. The separation of the flow chamber containing cells and the two upper and lower reservoirs is highlighted. Step 1 allows solution to flow from the upper reservoir of piece B to piece D of the chamber where cells are seeded. Step 2 allows solution to flow from piece D down to the lower reservoir, piece F. Please click here to view a larger version of this figure.
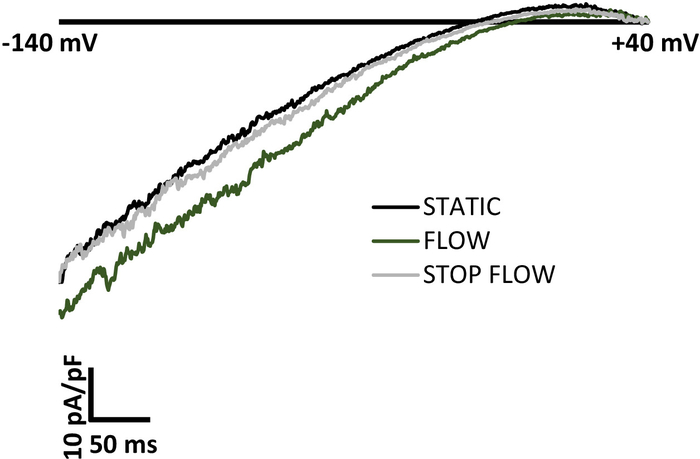
Figure 3: Representative traces of shear-induced current activation of Kir channels and return to baseline current levels upon removal of flow. A good positive control for mechanoactivation of ion channels is the return to baseline current levels initially observed in a static bath upon cessation of the mechanical stimulus. Inwardly rectifying K+ (Kir) channel currents are activated within seconds by shear stress when gravity solution is allowed to flow to the MPP flow chamber. Upon cessation of flow to the chamber, Kir currents quickly return to baseline static levels observed prior to flow. A ramp of -140 mV to +40 mV was applied to the patch over 400 ms. The bath solution contained 60 mM K+ and the reversal potential was ~-20 mV. The representative traces were generated from an endothelial cell freshly isolated from mouse mesenteric arteries. Please click here to view a larger version of this figure.
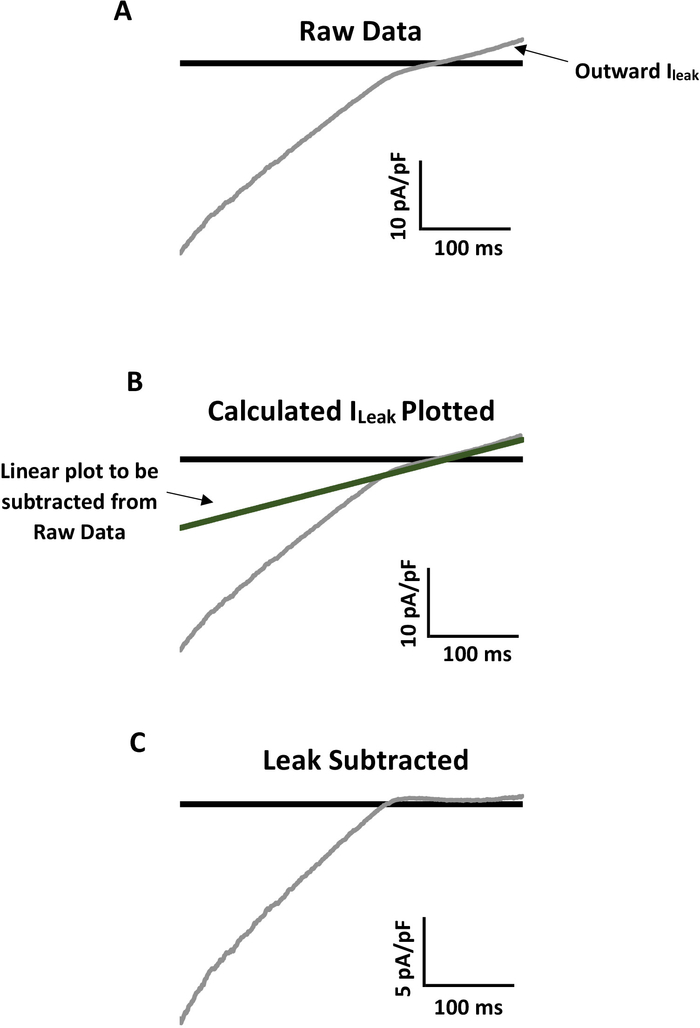
Figure 4: Example of leak subtraction for accurate analysis of mechanoactivated Kir current. (A) Representative raw recording of Kir current from a primary mouse mesenteric endothelial cell with notable linear outward leak current (Ileak). A ramp of -140 mV to +40 mV was applied to the patch over 400 ms. The bath solution contained 60 mM K+ and the reversal potential was ~-20 mV. (B) Ileak prevents analysis of real Kir channel activity. To subtract Ileak, first calculate the linear slope conductance of Ileak (Gslope = (Ia-Ib)/(Va-Vb)). Multiply Gslope by corresponding voltages of the entire raw trace to plot Ileak on the raw data. The line should overlay the outward linear leak exactly. (C) Subtract the plotted Ileak from the entire trace so that the linear outward current is ~0 pA/pF and the real Kir current can be analyzed. Notice in panel C that the inward Kir current is approximately half that of the original raw data trace in panel A. Please click here to view a larger version of this figure.
| Height (mm) | Width (mm) | Length (mm) | Additional information | ||
| Piece A | 8 | 80 | 98 | Contains a 3 mm diameter hole for inlet perfusion tubing (see Table of Materials) and 53 mm x 60 mm rectangular space for access to middle pieces and cells | |
| Piece B | 1 | 80 | 98 | Contains a space (20 mm diagonal) underneath perfusion hole of Piece A and three 2 mm x 17 mm slits for access to cells | |
| Piece C | 1 | 80 | 98 | Contains a 22 mm x 50 mm space where Piece D is adhered using the silicone elastomer solution (see Table of Materials) | |
| Piece D | 0.16 | 24 | 40 | Rectangular glass slide bottom of the flow chamber | |
| Piece E | 5 | 80 | 120 | Contains a 45 mm x 50 mm space to allow visualization of cells on Piece D. A 20 mm x 45 mm space is present for the reservoir vacuum outlet, Piece F, to be adhered | |
| Piece F | 0.16 | 24 | 50 | Rectangular glass slide bottom of vacuum outlet reservoir | |
| Assembled MPP | 15 | 80 | 120 | Flow chamber is separated from inlet perfusion and outlet vacuum by two steps to avoid disruption of well-defined laminar shear | |
| Flow Chamber of Assembled MPP | 1 | 22 | 42 | Piece D is the glass slide bottom of the flow chamber | |
Table 1: Dimensions of the MPP (assembled and disassembled) and additional information specific to each part of the device.
| Solution | Recipe (in mM) | pH | ||
| Dissociation | 55 NaCl, 80 Na-glutamate, 6 KCl, 2 MgCl2, 0.1 CaCl2, 10 glucose, 10 HEPES | 7.3 | ||
| (Cell isolation) | ||||
| Bath (Electrophysiology) | 80 NaCl, 60 KCl, 1 MgCl2, 2 CaCl2, 10 glucose, 10 HEPES | 7.4 | ||
| Pipette (Electrophysiology) | 5 NaCl, 135 KCl, 5 EGTA, 1 MgCl2, 5 glucose, 10 HEPES | 7.2 | ||
Table 2: Examples of solutions with recipes used in the experiments.
| Endothelial cell isolation protocol | References | |
| Cocktail of neutral protease and elastase (0.5 mg/mL each; 1 h at 37 °C) followed by collagenase type I (0.5 mg/mL; 2.5 min) | 6,7 | |
| Gentle mechanical scraping of a 5-cm2 region located at the inner wall of the descending thoracic aorta | 11 | |
| NA | 17 | |
| NA | 3 | |
| Collagenase type IA (1 mg/mL) for 14 min at 37 °C | 8 | |
Table 3: Methodology to study mechanosensitive ion channels using electrophysiological techniques.
