Example experiment I was run in Římov water reservoir (South Bohemia, CZ), which is a natural site with lower natural in situ predator and prey abundance. Representative data is reported for the omnivorous ciliate species Halteria grandinella, which is an abundant and efficient grazer of picoplankton (<2 µm) particles10,16,17,18,22. Figure 3 shows box-and-whisker plots of numbers of FLB per cell of Halteria sp. from the Římov reservoir (Figure 3A), which was recalculated to rates of bacterial uptake per hour (Figure 3B) detected in four individual experiments conducted in April, May, August, and September. There was high variability in ciliate uptake rates, largely caused by the temporal differences in water temperature.
It should be noted that the Q10 parameter reflects the fact the microbial processes run approximately 2.5x faster with a temperature increase of 10 °C12, which also holds for ciliate uptake rates on bacteria. With this physiological rule in mind, considerably different proportions of FLB and incubation times were used for different seasons (for details, see Figure 3A). Thus, the expected temperature effect was compensated for, and the experimental setting yielded optimized average and median values of uptake rates approximately between 5-10 FLB per ciliate cell. Generally, these amounts of ingested FLB are easily countable (see examples in Figure 2, two left photographs), generating precise estimates of the tracer (mostly being between 1-15 FLB per ciliate) uptake rates. However, due to modified FLB tracer added (%) and different times of sample incubation the absolute values (expressed as number of bacteria grazed ciliate per hour) differed significantly (p < 0.01, Kruskal-Wallis test; followed by Dunn's multiple-comparison test, p < 0.05; see examples in Figure 3B) among the experiments. The data also illustrate typical natural variability in absolute bacterivory rates in the planktonic populations of Halteria grandinella, with a close match of their mean and medium values (Figure 3).
In the presence of highly efficient bacterivorous ciliates in samples, such as peritrichous ciliates, they can become heavily "over-labeled" by FLB in typical tracer amounts of 5%-10% of total bacteria (see right side photograph in Figure 2). This may strongly limit the accurate quantification of ingested FLB. In such cases, it is suggested to run additional parallel incubations with only low amounts of FLB accounting for only 1.5%-3% of total bacteria. However, generally both the tracer amounts as well as incubation times can be manipulated to optimize the number of FLB per cell (Figure 2).
Example experiment II: Displayed is the data from a system with large predator and prey abundances, where only an extremely small sample volume is available to experimentally estimate bacterivory rates of the ciliate Tetrahymena utriculariae25. It is a moderate bacterial grazer living in high abundance exclusively in traps of carnivorous Utricularia reflexa plants26,27. Figure 4 shows box-and-whisker plots of number of FLB per cell of T. utriculariae under different experimental settings (Figure 4A,B) that is recalculated into rates of bacterial uptake per hour (Figure 4C,D) detected in young, mature, and old traps. Interestingly in traps, chloroplast-bearing populations of the ciliate T. utriculariae were detected, while apochloric populations of T. utriculariae were isolated from traps and maintained on mixed bacterial suspension growing on wheat grains in the dark (for details, see Figure 1 in a previous publication26).
The chloroplast-bearing populations live in light-illuminated traps; thus, the chloroplasts can provide an additional organic carbon source and oxygen to the ciliate host. One of the hypotheses tested was that the apochloric ciliate populations grazed bacteria significantly faster, as the bacteria represent the only particulate source of organic carbon available to dark-grown isolated subpopulations of the ciliate.
Indeed, while there were no significant differences in bacterivory rates of the ciliates living in young, mature and old traps of Utricularia reflexa (Figure 4A,C), the apochloric populations of T. utriculariae grazed bacteria significantly (p < 0.01, Kruskal-Wallis test; followed by Dunn's multiple-comparison test, p < 0.05), approximately 3x faster than the chloroplast-bearing ciliates living in young, mature, and old traps (Figure 4C,D). Note that again, both the tracer amounts as well as incubation times (Figure 4A,B, top) were modified to optimize number of FLB per cell (generally between 1-15), with average and median values around 5 FLB/ciliate. These numbers are distinguishable in ciliate food vacuoles and allowed accurate tracer counting. However, expressed in absolute numbers of bacteria grazed per hour, the chloroplast-bearing and apochloric populations grazed approximately 350 and 1,000 bacteria ciliate per hour, respectively. This experimental set-up brought new insights into the metabolic and physiologic traits of two distinct subpopulations of the same ciliate species living under strikingly different environmental constraints25,26,27.
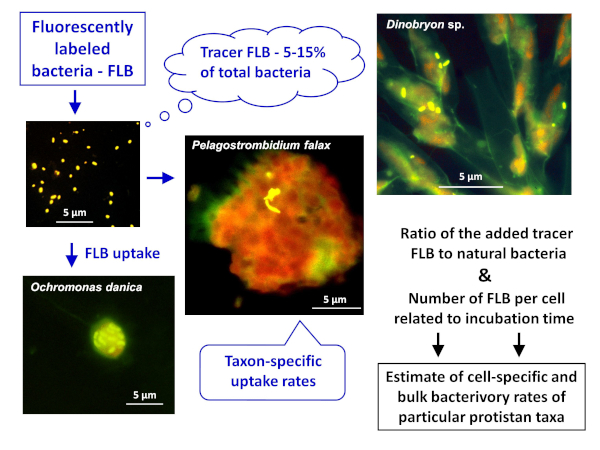
Figure 1: Workflow of using fluorescently labeled bacteria (FLB) to estimate cell- and species-specific grazing rates from the ratio of ingested tracer FLB to total number of natural bacteria in the sample. For more details, see section 7 of the protocol. Please click here to view a larger version of this figure.
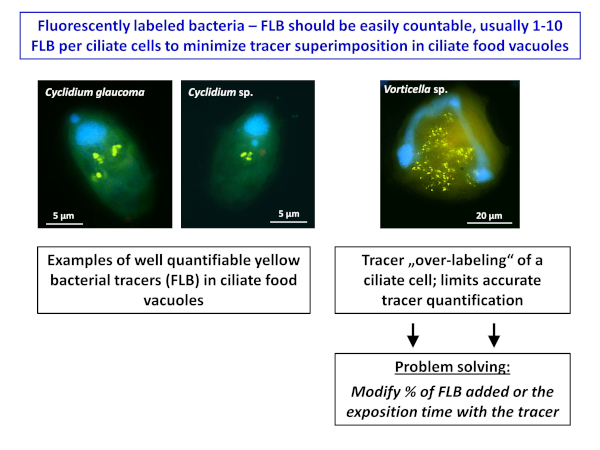
Figure 2: Examples of ciliate cells from plankton of a eutrophic fishpond. Examples are shown from the pond with countable FLB in ciliate cells (generally 1-10 tracer FLB per cell, the left two microphotograps) compared to a peritrichous ciliate Pelagovorticella natans (the right side microphotograph). Even during a short, 5 min incubation period, it became "over-labeled" by the tracer FLB, making quantitation of the ingested FLB inaccurate or almost impossible. In this case, it is suggested to decrease the tracer amount to 1.5%-3% of total bacteria. However, generally both the tracer amounts and incubation times can be manipulated to optimize the number of FLB ingested per cell. For more details, see section 7 of the protocol. Please click here to view a larger version of this figure.
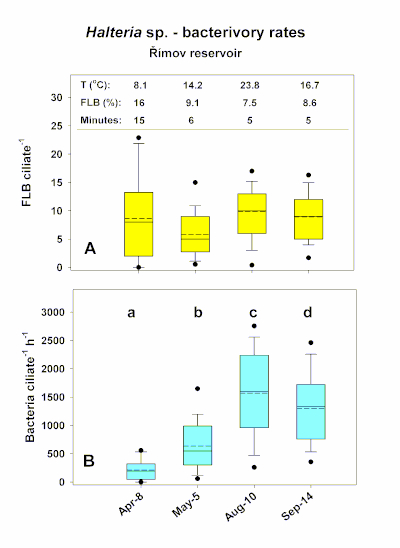
Figure 3: Box-and-whisker plots of numbers of FLB per cell of Halteria sp. from the Římov reservoir (Exp I) (A), recalculated to rates of bacterial uptake per hour (B). The data were detected under different seasonal settings, represented by four examples from April to September. The top of panel A shows information on water temperature, different FLB tracers added (%), and different times of sample incubation. It should be noted that the latter two parameters can be modified to optimize number of FLB per cells, with average (full line) and median (dashed line) values approximately between 5-10 FLB per ciliate cell (A). The bars show the 25th and 75th percentiles of all data (50-180 cells inspected) and whiskers stand for the 1th and 99th percentiles. (B) Different small letters indicate significant differences in cell-specific bacterivory rates of Halteria sp. during the studied period. Please click here to view a larger version of this figure.
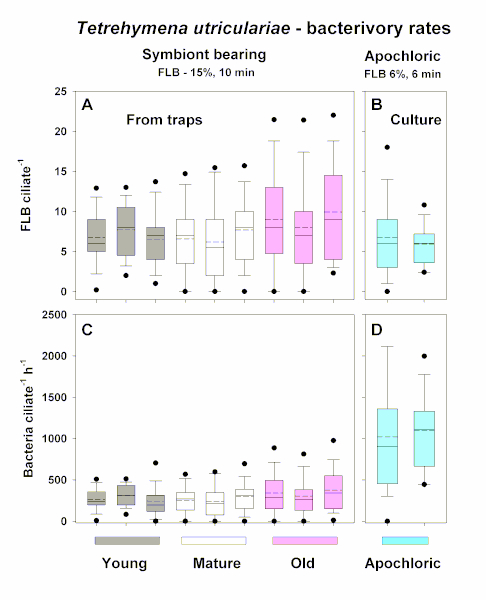
Figure 4: Box-and-whisker plots of numbers of FLB per cell. Plots are shown of chloroplast-bearing Tetrahymena utriculariae from triplicate treatments of young, mature, and old traps of Utricularia reflexa (Exp II) (A), recalculated to rates of bacterial uptake per hour (C). The data were compared to bacterial uptake rates of the duplicate apochloric populations of T. utriculariae (B,D) isolated from traps but maintained on mixed bacterial suspension growing on wheat grains in dark. On the top of panels A and B, different FLB tracers added (%) and different times of sample incubation are shown. It should be noted that the latter two parameters were modified to optimize number of FLB per cells, with average (full line) and median (dashed line) values approximately between 5-10 FLB per ciliate cell (A,B). The bars show the 5th and 95th percentiles of all data (50-100 cells inspected), and whiskers stand for the 1th and 99th percentiles. Please click here to view a larger version of this figure.
