The methods described here allow for the visualization of the integrity of the BBB throughout the CNS in D. melanogaster embryos and larvae (Figure 1). Upon completion of BBB formation in late embryogenesis, the BBB functions to exclude large molecules from the brain and VNC5. This protocol takes advantage of this function to assay BBB formation. When wild-type (Oregon R) late stage 17 (20−21 h old) embryos were injected with 10 kDa dextran conjugated to sulforhodamine 101 acid chloride fluorescent dye, the large dextran molecule was excluded from the VNC, as expected (Figure 2A). In order to demonstrate the effect of genetic mutations on BBB integrity, embryos with mutations in the raw gene were utilized. Previously, the raw gene has been demonstrated to regulate germ band retraction during embryogenesis, and more recently has been shown to function in glia to regulate morphological changes in the VNC during development28. Heterozygous raw1 mutant embryos exhibited an intact BBB, similar to the results observed in wild-type control embryos (Figure 2B). In contrast, homozygous raw1 mutant embryos exhibited defects in the integrity of the BBB, with 10 kDa dextran dye flooding into the VNC, indicating that the BBB failed to form (Figure 2C). These results demonstrate the ability of this technique to assay BBB formation during embryonic stages.
Previous studies have demonstrated that a defect in subperineurial glia polyploidization results in a disruption of the BBB that can be observed in third instar larval stages7,9. Thus, defects in BBB formation and/or maintenance could result in a compromised BBB during later stages of development, making it desirable to assay integrity of the barrier in the third instar larval stage. Therefore, the protocol utilized for assaying BBB integrity in embryonic stages has also been optimized for use in larvae. In Oregon R control samples, 10 kDa dextran fails to penetrate the BBB and is excluded from the brain and VNC (Figure 2D,E). The dye accumulates at the periphery of the BBB.
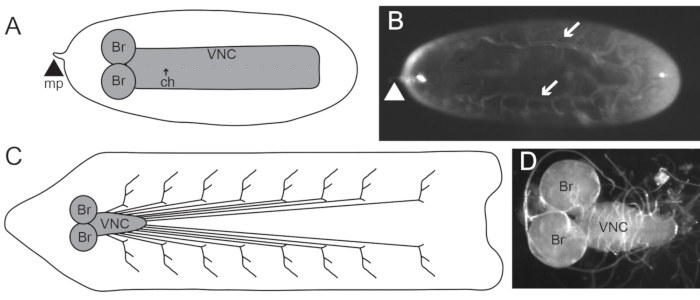
Figure 1: The nervous system of stage 17 embryos and third instar larva. (A) Schematic of a ventral view of a stage 17 embryonic central nervous system (CNS). The CNS consists of the brain (Br) and ventral nerve cord (VNC), which has dorsoventral channels (ch). The micropyle (mp; arrowhead) at the anterior end can be used to orient the embryo. Posterior to the right. (B) Stage 17 embryo oriented with the dorsal side up and posterior to the right, as recommended in step 1.1.10. Arrows are pointing to the trachea. Arrowhead indicates the mp. Posterior to the right. (C) Schematic of the nervous system in the third instar larva. The brain (Br) and VNC compose the CNS, while the nerves extending from the VNC synapse onto body wall muscles and are part of the PNS. Posterior to the right. (D) Dissected third instar larval brain and VNC. Posterior to the right. Please click here to view a larger version of this figure.
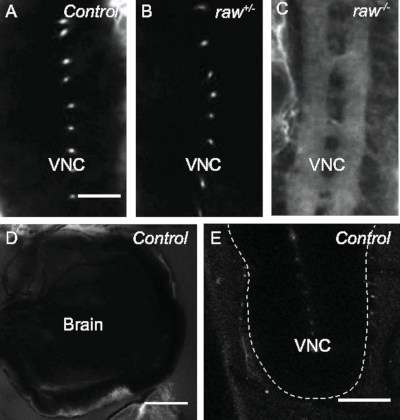
Figure 2: Assay for blood-brain barrier (BBB) formation. (A-C) Late stage 17 embryos (20−21 h old) injected posteriorly with 10 kDa sulforhodamine 101 acid chloride dextran. Posterior down. Scale bar = 20 μm. Dots seen in controls are dorsoventral channels that span the ventral nerve cord (VNC). (A) Oregon R control. Dye uptake in 6.25% of samples, n = 16. (B) raw1/+ sibling control. Dye uptake in 6.67% of samples, n = 15. (C) Homozygous raw1/1 mutant. Dye uptake in 100% of samples, n = 22. (D) Oregon R control third instar larval brain. Dye accumulates at the BBB, but does not penetrate into the CNS, n = 7. Scale bar = 50 μm. (E) Oregon R control third instar larval VNC. Dye accumulates at the BBB, but does not penetrate into the CNS, n = 11. Dashed line outlines the VNC. Scale bar = 50 μm. Please click here to view a larger version of this figure.
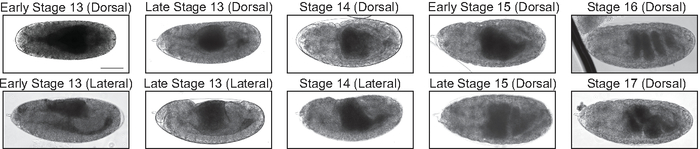
Figure 3: Midgut morphology in late embryonic development. Transmitted light images of stage 13−17 embryos allow for visualization of gut morphology (dark grey regions in the posterior half of the embryo). Midgut morphology can be used to determine the stage of embryonic development, which is helpful when determining if embryos are being aged appropriately for injection. Scale bar = 100 μm. Please click here to view a larger version of this figure.
| Solution | Recipe |
| 2% Agarose gel | Add 0.8 g of agarose to 40 mL of double-distilled H2O (ddH2O) in an Erlenmeyer flask and microwave to dissolve agar. Pour into a gel casting tray and allow the gel to solidify for 30 min at room temperature. |
| Apple juice agar plates | Measure 45 g of agar and 1.5 L of ddH2O into a 4 L flask. Autoclave for 40−45 min at 121 °C. Measure 50 g of sugar and 0.5 L of apple juice into a 1 L beaker and stir on low heat (setting 3) to dissolve sugar, taking care not to burn it. Following autoclaving, add the preheated sugar and apple juice to the agar and water. Stir on low with the heat off to allow to cool until you can touch it. Add 15 mL of 70% tegosept and stir to disperse. Pour into a 0.5 L beaker. Spray with ethanol to remove bubbles or flame with a Bunsen burner. Pour into 60 mm petri dishes. Allow to set for at least 24 h, or until there is minimal condensation on the lids of the petri dishes and store at 4 °C. |
| 50% Bleach | Add 15 mL of ddH2O and 15 mL of household bleach into a conical tube. |
| 80% Glycerol | For 10 mL, add 8 mL of autoclaved ddH2O and 2 mL of glycerol to a 15 mL conical tube. Incubate on a rocker until solution is homogeneous. |
| 1x PBS | For 1 L, dilute 100 mL of 10x PBS in 900 mL of ddH2O. |
| 10x PBS | For 2 L, dissolve the following in 1,600 mL of ddH2O: 160 g of NaCl, 4 g of KCl, 28.8 g of Na2HPO4, and 4.8 g of KH2PO4. Adjust pH to 7.5 with HCl. |
| PBTx (PBS + 0.1% nonionic surfactant) | For 1 L, combine 100 mL of 10x PBS, 10 mL of 10% nonionic surfactant, and 890 mL of ddH2O. |
| 70% Tegosept | Mix 1 g of p-hydroxybenzoic acid, methyl ester for every 10 mL of 100% ethanol. Store at -20 °C. |
| 10% Nonionic surfactant | For 50 mL, add 45 mL of autoclaved ddH2O and 5 mL of nonionic surfactant to a conical tube. Rotate on the rocker until the solution is homogeneous. |
| Yeast paste | Mix dry active yeast with ddH2O in a 50 mL plastic beaker until smooth. |
Table 1: Reagents and buffers used throughout this protocol.
| Genotype | Stock |
| w[1118]; ln(2LR)Gla, wg[Gla-1]/CyO, P{w[+mC]=GAL4-twi.G}2.2, P{w[+mC]=UAS-2xEGFP}AH2.2 | Bloomington #6662 |
| w{*]; sna[Sco]/CyO, P{w[+mC]=Dfd-EYFP}2 | Bloomington #8578 |
| w{*]; L[2] Pin[1]/CyO, P{w[+mC]=GAL4-Kr.C}DC3, P{w[+mC]=UAS=GFP.S65T}DC7 | Bloomington #5194 |
| Df(1)JA27/FM7c, P{w[+mC]=GAL4-Kr.C}DC1, P{w[+mC]=UAS-GFP.S65T}DC5, sn[+] | Bloomington #5193 |
| w[*]; ry[506] Dr[1]/TM6B, P{w[+mc]=Dfd-EYFP}3, Sb[1] Tb[1] ca[1] | Bloomington #8704 |
| y[1] w[*]; D[*] gl[3]/TM3, P{w[+mC]=GAL4-Kr.C}DC2, P{w[+mC]=UAS-GFP.S65T}DC10, Sb[1] | Bloomington #5195 |
| Oregon R | Multiple Strains Available |
| raw[1] cn[1] bw[1] sp[1]/CyO | Bloomington #2749 |
| P{w[+mW.hs]=GawB}elav[C155] | Bloomington #458 |
| w[1118]; P{w[+m*]=GAL4}repo/TM3, Sb[1] | Bloomington #7415 |
| P{UAS-GFP} | Multiple strains available |
Table 2: Fly strains. Fly strains discussed throughout this protocol. The Bloomington Drosophila Stock Center stock number is provided where applicable.
| Cycle | Heat | Pull | Velocity | Time | Pressure | Ramp |
| 1 | 590 | 115 | 15 | 250 | 600 | X |
| 2 | 575 | 130 | 60 | 250 | 600 | X |
Table 3: Micropipette puller settings. Micropipette puller settings used to generate needles for injection in this protocol.
