This protocol describes a method to isolate and sort microglia from different brain regions in one adult perfused brain hemisphere, followed by scRNA-seq. We use douncing to create single cell suspension and also as a first step to enrich microglia. Insufficient or over-douncing reduces the yield. In addition, adult mouse brains contain high levels of myelin, which can also reduce sorting efficiency and yield if not removed properly. Therefore, we examine the cell suspension under microscope by using trypan blue and a hemocytometer to estimate the yield, cell viability and efficacy of myelin removal (step 4.9) before performing antibody staining (Figure 1). Total cell counts at this point should be over 30,000 for cortex, and over 5,000 for other tissues. Over 90% of cells should be viable with little myelin debris.
We use a FACS machine to sort microglia (or myeloid cells), which are typically CD45 low and CD11b positive. At least for the cortical tissue, successful isolation should generate over 80% microglia out of all live single cells (Figure 2). The dying/dead population represents only a small fraction of the preparation (around 10%).
Once individual microglia are captured into the lysis buffer, RNA is released and subsequently reverse transcribed to cDNA, which is then amplified for 23 cycles. It is important to check the quality of these cDNA samples—at least a portion of them—before making libraries. As a capillary electrophoresis platform, the fragment analyzer and high-sensitive NGS fragment kits (1−6,000 bp) provide quick and accurate information about size distribution as well as quantity of cDNA molecules present in each well of a 96-well plate (Figure 3A). Samples showing a smear (500−5,000 bp) and above certain concentration threshold (e.g., 0.05 ng/µL) can be used to make libraries. Similarly, the pooled libraries should be tested on a bioanalyzer before sequencing (Figure 3B).
We sequence the samples to a depth of over 1 million raw reads per cell, which saturates the detection power of this scRNA-seq methodology16. With about 60% mapping rate, over 2,000 genes per microglial cell can be detected. We obtained published data that were generated using this isolation method17, and demonstrate its reproducibility from independent experiments and sensitivity for detecting microglia-specific genes across the sequenced population (Figure 4).
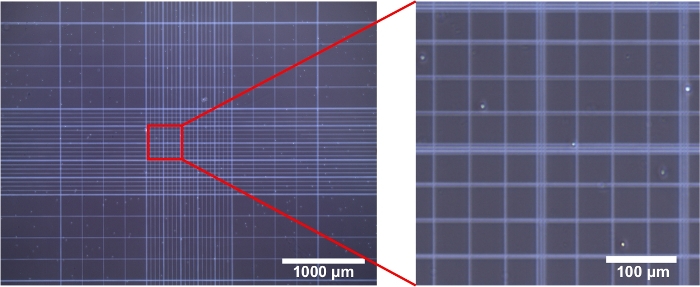
Figure 1: Estimation of microglia isolation yield, cell viability and efficiency of myelin removal. The left panel showed the bright field image (4x magnification) of trypan blue stained cells (from cortex) after passing through myelin removal columns. Results for other regions would look similar but with fewer cells. The vast majority (if not all) of cells appeared bright (non-stained) and round due to loss of processes. The right image was the zoom-in (20x) of the boxed area and little myelin debris was present. Please click here to view a larger version of this figure.
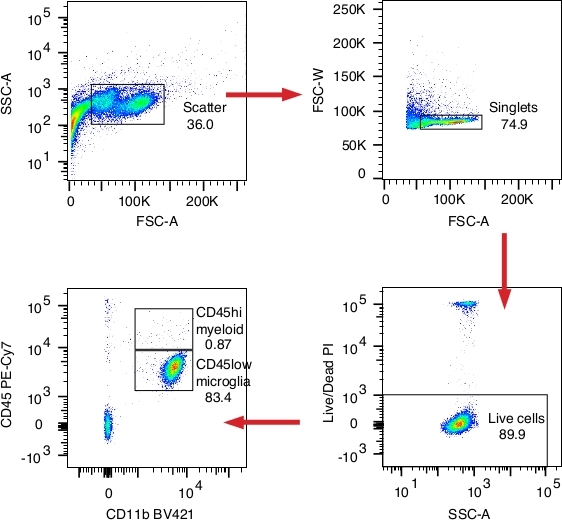
Figure 2: Gates used for sorting microglia. Data showed the gating strategy for sorting microglia from the cortex, and the same strategy was used for other regions. Cell debris was first excluded from the scatter plot, and single cells were gated by forward scatter-area (FSC-A)/forward scatter-width (FSC-W). Live cells were gated by PI negative staining, which were from about 90% of all single cells. Microglia, representing roughly 80% of live cells, were sorted from the CD45 low CD11b+ gate. A total of ~30,000 microglia could be isolated and sorted from the cortex tissue (from one hemisphere of a 3-month old mouse). Index sorting was performed on a FACS machine. Please click here to view a larger version of this figure.
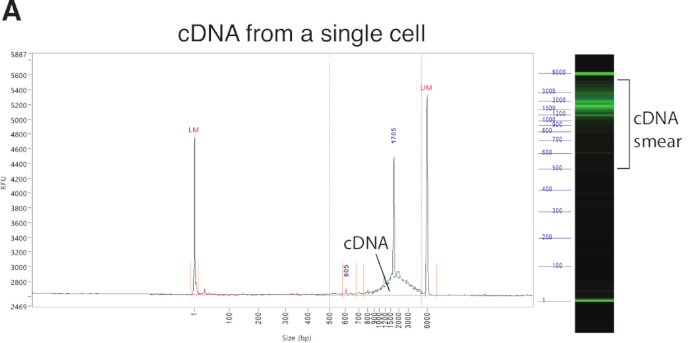
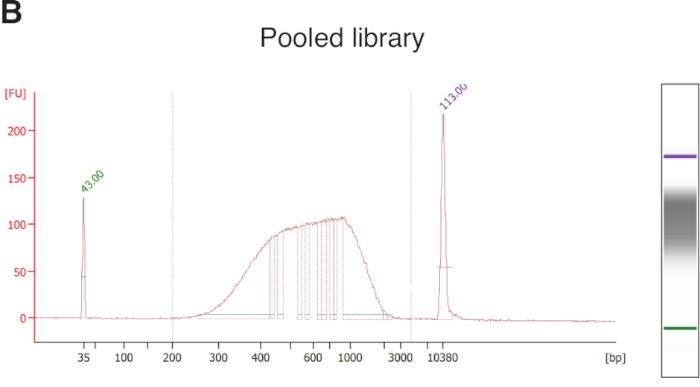
Figure 3: Representative quality control results for amplified cDNA from a single microglial cell and an eventual pooled library. (A) Using a fragment analyzer, all fragments are plotted with their sizes on the X axis, and relative fluorescence intensity on the Y axis, signifying the abundance of a given sized cDNA. LM (lower marker, 1 bp) and UM (upper marker, 6,000 bp) are loading markers with known concentrations used for measuring cDNA quantity. Successfully amplified cDNA from a representative microglial cell forms a curve (green dotted line) on the size distribution graph or a smear (labeled bracket) on the gel graph between 500 bp and 5,000 bp. Other labeled peaks were amplified from ERCC spike-in molecules or ribosomal RNA. The concentration of cDNA between 500 bp and 5,000 bp can be quantified, and those samples with concentrations higher than 0.05 ng/μL and showing such typical curves are retained for library preparation. RFU, relative fluorescence unit. (B) Representative quality control result on a bioanalyzer showing size distribution of a final pooled library (with about 380 cells). Typically, it ranges from 200 bp to 2,000 bp with an average size of 400−600 bp. The two sharp peaks were loading markers. FU, fluorescence unit. Please click here to view a larger version of this figure.
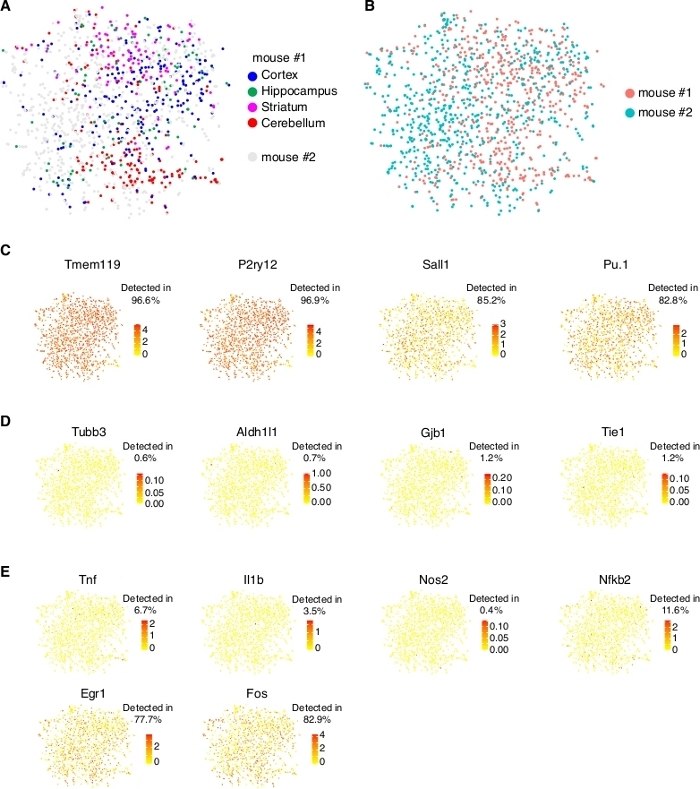
Figure 4: scRNA-seq analysis of microglia isolated from 4 brain regions of two male mice. (A) tSNE plot showing intermingled pattern of regional microglia (only cells from mouse #1 were highlighted). They do not form distinct clusters according to region origins beyond subtle shift into concentrated areas on the tSNE plot (possibly due to batch effects). This observation is consistent with limited regional heterogeneity of microglia based on global gene expression in the adult mice (see a recent publication2 for further discussion on this topic). (B) tSNE plot showing overlapping pattern of microglia from two individual mice that were processed independently. Although small batch effects may exist (cells concentrating in certain areas of the plot), these cells do not form distinct clusters according to animal origins. This result suggests the reproducibility of the protocol for comparing data between experiments. (C) Expression of microglia signature genes detected by scRNA-seq showing over 95% detection rate for specific markers, such as Tmem119 and P2ry12, and over 80% detection rate for known transcription factors such as Sall1 and Pu.1. The data were re-analyzed from published literature17. (D) Vast majority of isolated microglia lack expression of genes specific to other cell types, such as Tubb3 (neurons), Aldh1l1 (astrocytes), Gjb1 (oligodendrocytes), and Tie1 (endothelial cells). (E) Expression of genes related microglial activation or stress. Classical markers, Tnf, Il1b, Nos2, and Nfkb2, which are lowly expressed, are shown on the top. Early response genes, Egr1 and Fos, are shown at the bottom (see discussion). Please click here to view a larger version of this figure.
| Reagent | Volume (µL) |
| Cell lysis | 4 |
| Reverse transcriptase (100 U/µL) | 0.95 |
| Rnase inhibitor | 0.25 |
| 5x First strand buffer | 2 |
| Dithiothreitol (DTT; 100 mM) | 0.5 |
| Betaine (5 M) | 2 |
| MgCl2 (1 M) | 0.06 |
| Template switch oligo (TSO; 100 µM) | 0.1 |
| H2O | 0.14 |
| Total | 10 |
Table 1: Reverse transcription condition. Reagent volumes for one reverse transcription reaction are provided.
| Reagent | Volume (µL) |
| Reverse transcription product | 10 |
| 2x PCR master mix | 12.5 |
| ISPCR primer (10 µM) | 0.25 |
| Lambda exonuclease | 0.1125 |
| H2O | 2.1375 |
| Total | 25 |
Table 2: PCR amplification condition. Reagent volumes for one PCR amplification of cDNA from a single cell are provided.
| Plate-based (this protocol) | Droplet-based (10x Genomics) | |
| Sensitivity | More genes detected | Fewer genes detected |
| Full length | Yes | No (5' or 3' end) |
| Flexibility for cell numbers/populations | Suitable for characterization of small or rare subpopulations | Suitable for broad categorization of large cell populations |
| Throughput | Up to several thousands of cells | Hundreds to tens of thousands of cells |
| Unique molecular identifier | No | Yes |
| Cost per cell | $1−$5 | less than $1 |
| Experimental difficulty | More steps and usually require liquid handling robotics | Simple to perform with commercialized machines |
| Cell populations | Targeted by FACS sorting | Unbiased |
Table 3: Comparison between plate-based and droplet-based scRNA-seq methods. In this article, the plate-based full-length scRNA-seq procedure is provided, which has the advantages of higher sensitivity, full-length sequencing and flexibility for small numbers of cells as inputs. Droplet-based methods offer the advantages of higher throughput, lower cost, easy to perform and data in unique molecular identifiers. They are complementary depending on the purpose of the experiments.
