Characterization of mouse morbidity in influenza virus infection
After influenza virus infection, mice are less active and anorexic due to illness, which is reflected by severe weight loss, a commonly used symptom to monitor the mouse morbidity19. As shown in Figure 1a, PR8 virus-infected mice started to lose weight on day 6, reached the highest loss level on day 8 and returned to the initial level on day 10. As expected, weight loss was not observed all through the period in PBS-treated control mice. For in vivo symptoms, virus infection leads to robust lymphocytes expansion in the draining lymph node, mLN in this case. Therefore, significantly larger size of mLNs were observed in PR8 virus-infected mice than in control mice (Figure 1b). Taken together, these mice all showed expected symptoms and were qualified for the subsequent Tfh-associated immune response study.
Detection of Tfh differentiation and function-associated molecules
To analyze Tfh differentiation, mice were sacrificed on day 5, 7, 10 and 14 after infection and mLNs or spleens were isolated for flow cytometry analysis. Figure 2a and Figure 2b show the Tfh population gating strategy, with Tfh gated as PD-1hi CXCR5hi cells and non-Tfh as PD-1lowCXCR5low cells. With this gating strategy, the kinetics of Tfh differentiation during influenza virus infection were assayed. As shown in Figure 2c, Tfh differentiation initialized at day 5 and peaked at day 10. So we took samples of day 10 for further analysis. As shown in Figure 3a, robust Tfh cell differentiation was induced in influenza virus-infected mice compared with control mice. To analyze Influenza virus-specific Tfh cells, fluorochrome-labeled IAbNP311–325 MHC class II tetramers (NP311-325) were added in the polyclonal Tfh cells staining panel (Table 1). Both in mLNs and spleens from influenza virus-infected mice, NP311-325-specific CD4+ T cells were significantly induced and NP311-325-specific Tfh cells could be analyzed by addition of PD-1 and CXCR5 into analysis(Figure 3e). Because of essential roles of Bcl6 in Tfh differentiation, Bcl6+CXCR5+ cells can also represent the Tfh population. Consistently, Tfh cells identified with this strategy were also induced robustly (Figure 3b). We further analyzed expression of Bcl6 in Tfh and non-Tfh cells. As shown in Figure 3c, higher expression of Bcl6 in Tfh cells than that in non-Tfh cells indicates successful Bcl6 staining. With similar strategy, ICOS, another Tfh-associated molecule was also analyzed (Figure 3d). Due to the specialized role of Tfh cells in providing help for B cells, assay of the expression of IL21, which is secreted mainly by Tfh cells and demonstrated to directly regulate B cells survival and proliferation, could reveal Tfh cells function to some extent. As shown in Figure 3f, intracellular staining of IL21 revealed that PR8 infection induced significantly higher production of this cytokine, with unstimulated cells as gating control. Taken together, these assays could reflect basic information of Tfh differentiation and provide the insights into the B cell-help ability.
Detection of GC B and plasma cells development and influenza virus-specific antibodies in serum
The main function of Tfh cells is to provide B cell help in GCs, in which antibody class switching and affinity maturation occur. So GC B development could indirectly reflect differentiation and function of Tfh cells. GC B cells could be gated as B220+PNA+Fas+ cells (Figure 2d). Through this gating strategy, we assayed the kinetics of GC B cell response and found that GC B response started at day 10 and continued to increase at day 14 (Figure 2e). Comparison between PR8 virus-infected and control mice showed robust GC B were induced both in mLN and spleen after influenza virus infection(Figure 4a), which is consistent with the induced Tfh differentiation in PRB virus-infected mice. In addition, Immunofluorescence staining with IgD and PNA provides visualized images indicating induced GC reaction (green areas) in PR8 virus-infected mice (Figure 4d). Plasma cells, identified as IgDlowCD138+ cells(Figure 2d), were also generated in PR8 virus-infected mice (Figure 4b). Previous studies have identified that IFNƳ and IL21 could be secreted from both Th1 and Tfh cells in virus infection and induce IgG2 and IgG1 class switching, respectively20. Figure 4c depicts the generation of influenza virus-specific antibody by ELISA assay of HA-specific IgM, total IgG, IgG1, IgG2b and IgG2C. Together, all of these assays reflect the Tfh-associated B cell responses in influenza virus infection.
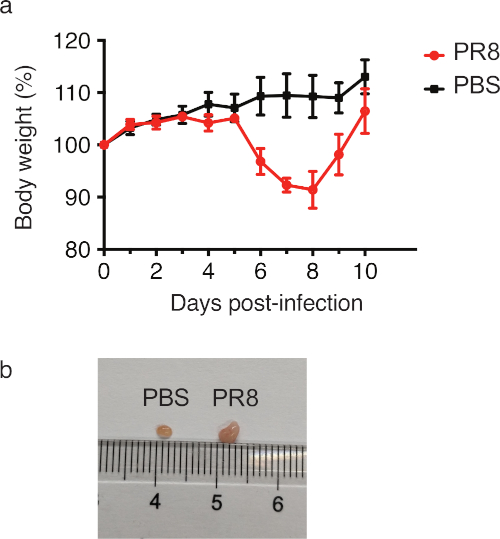
Figure 1: Characterization of mouse morbidity. 8-week-old male mice were infected with 40 PFU of PR8 influenza virus by intranasal inoculation. Mice were weighed daily for 10 days (a) and mLNs were isolated on d.p.i 10 (b). The error bars in (a) represent the mean ± SD. n = 4 mice per group. Please click here to view a larger version of this figure.
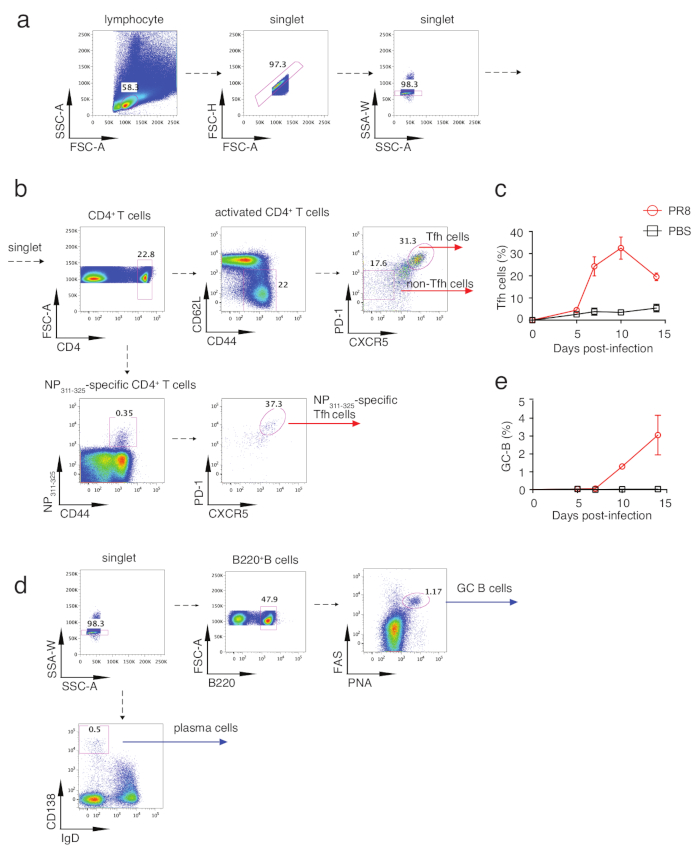
Figure 2: Gating strategy of Tfh cells and GC B cells. (a) Lymphocytes are defined by FSC-A and SSC-A, and cell singlets are gated with FSC-A, FSC-H and SSC-A, SSC-W. (b) After gating in CD4+ T cells, surface markers CD62L and CD44 are used to distinguish the naïve T cells (CD44loCD62Lhi) and activated T cells (CD44hiCD62Llo). Polyclonal Tfh cells can be gated from activated T cells as PD-1hi CXCR5hi population, conversely, non-Tfh cells as PD-1lowCXCR5low. PR8 virus-specific Tfh cells are defined as CD4+CD44+ NP311-325 tetramer+PD-1hi CXCR5hi cells. (c,e) Kinetics of Tfh frequency in activated cells (c) and GC B frequency in B220+ cells (e). (d) GC B cells are gated as B220+ PNA+FAS+ cells, and plasma cells are IgD–CD138+ cells. Please click here to view a larger version of this figure.
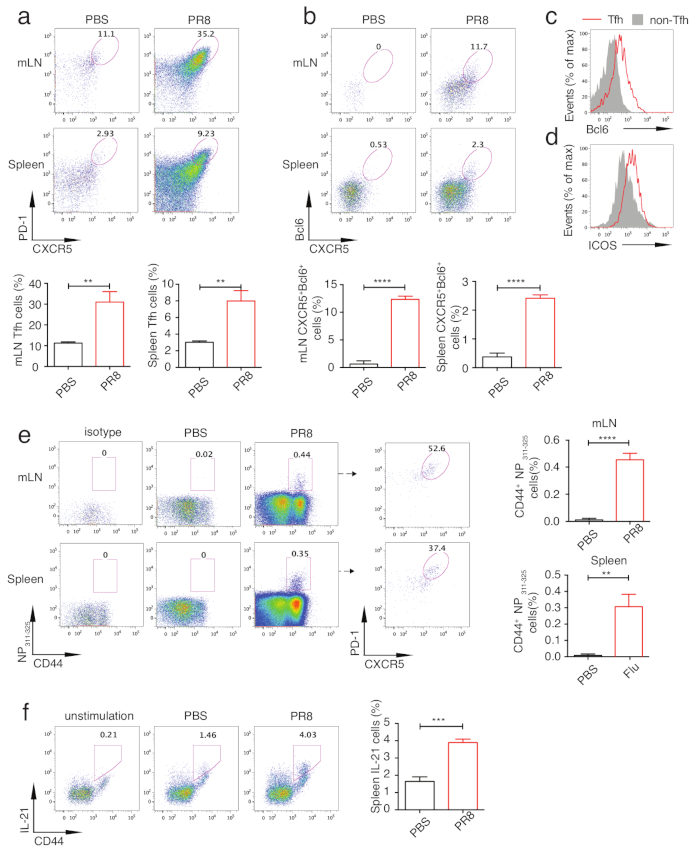
Figure 3: Analysis of Tfh differentiation in PR8 virus-infected mice. Mice were sacrificed on d.p.i 10 and mLNs and spleens were isolated for Tfh differentiation analysis. (a) Tfh percentage in mLNs and spleens in PR8 virus-infected mice and PBS-treated mice (upper panel). The statistics of Tfh cells (lower panel). (b) The intracellular staining of Bcl6 in CD4+CD44hi T cells (upper panel). The statistics of Bcl6+CXCR5+ cells (lower panel). (c) Bcl6 and (d) ICOS expression in Tfh (line-red) and non-Tfh cells (solid-gray). (e) Gating of NP311-325-specific CD4+ T cells in mLNs and spleens of PR8 virus-infected and PBS-treated mice (left panel). The percentage of PR8 virus-specific Tfh cells in mLNs and spleens (middle panel). “Isotype” indicates staining with irrelevant tetramer control. The statistics of NP311-325-specific CD4+ T cells (right panel). (f) Intracellular staining of IL-21 in splenic CD4+ T cells from PR8 virus-infected and PBS-treated mice, the unstimulation shown as control (left). The statistics of IL-21 staining (right). **P < 0.01, ***P < 0.001 and **** P < 0.0001 (two-tailed Student’s t-test). The error bars represent the mean ± SD. n = 3 mice per group. Please click here to view a larger version of this figure.
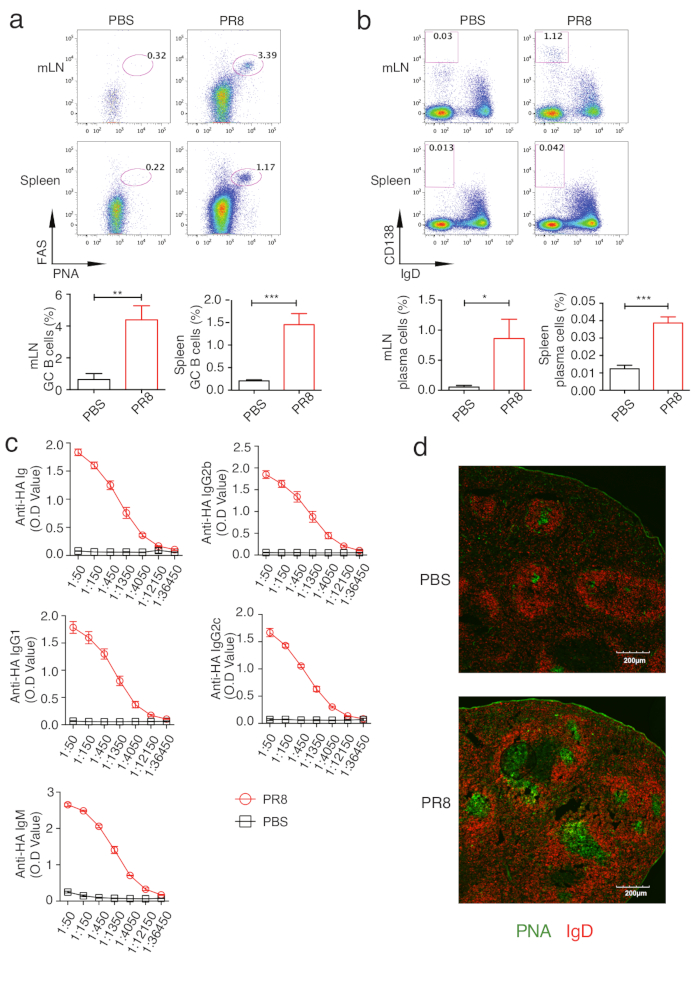
Figure 4: Analysis of GC B cell-associated response in PR8 virus-infected mice. Mice were sacrificed on d.p.i 10 and the mLNs and spleens were isolated for analysis. (a) The percentage of GC B cells (upper panel). The statistics of GC B cells (lower panel). (b) The percentage of plasma cells (upper panel). The statistics of plasma cells (lower panel). (c) Quantification of PR8 virus HA-specific IgG, IgM, IgG1, IgG2b and IgG2c in the serum (d.p.i 14) of PR8 virus-infected mice and PBS-treated mice. (d) Confocal microscopy of B cell follicles (IgD+, Red) and GCs (PNA+, Green) in the spleen samples of PR8 virus-infected mice and PBS-treated mice (d.p.i 10). *P < 0.5, **P < 0.01, and ***P < 0.001 (two-tailed Student's t-test). The error bars represent the mean ± SD. n = 3 mice per group. Please click here to view a larger version of this figure.
| surface marker | fluorochrome | clone | volume per sample(ul) |
| CD4 | Percp-eFluor 710 | GK1.5 | 0.2 |
| CD44 | eVolve 605 | IM7 | 0.2 |
| CD62L | FITC | MEL-14 | 0.2 |
| ICOS | BV421 | 7E.17G9 | 0.2 |
| PD1 | PE/Cy7 | 29F.1A12 | 0.3 |
| Streptavidin | PE | 0.2 |
Table 1: Surface marker (except for CXCR5) antibodies panel for staining Tfh cells (PD-1hiCXCR5hi).
| surface marker | fluorochrome | clone | volume per sample(ul) |
| CD4 | Percp-eFluor 710 | GK1.5 | 0.2 |
| CD44 | FITC | IM7 | 0.2 |
| PD1 | PE/Cy7 | 29F.1A12 | 0.3 |
| Streptavidin | BV421 | 0.5 |
Table 2: Surface marker antibodies (except for CXCR5) panel for staining Bcl6 in Tfh cells.
| surface marker | fluorochrome | clone | volume per sample(ul) |
| CD4 | Percp-eFluor 710 | GK1.5 | 0.2 |
| CD44 | FITC | IM7 | 0.2 |
Table 3: Surface marker antibodies panel for intracellular staining of IL21.
| surface marker | fluorochrome | clone | volume per sample(ul) |
| B220 | APC | RA3-6B2 | 0.2 |
| IgD | eFluor 450 | 11-26c | 0.2 |
| CD95 | PE/Cy7 | Jo2 | 0.3 |
| PNA | FITC | 0.3 | |
| CD138 | PE | 281-2 | 0.2 |
Table 4: Surface marker antibodies panel for staining GC B and plasma B cells.
