The gating strategy for the analysis of stem cell purity is depicted in Figure 1. Figure 1A–C shows the purified CD34+ population and Figure 1D–F the CD34- flow-through used to illustrate that the minimal amount of the CD34+ population is lost in the isolation process. The purity of isolated CD34+ stem cells was between 85%–95% with less than 1% T-cell contamination. Figure 1G depicts CCR5 bands from one adult human control donor with the CCR5Δ32/wt genotype, followed by bands from two CCR5wt/wt and one CCR5Δ32/wt stem cell donors. The frequency of the genotype CCR5Δ32/wt in a group of 19 donors was 15.8% (Figure 1H). This is in agreement with larger epidemiological studies14,15 reporting the genotype in up to 23.6% of investigated persons in Denmark.
Human CD45+ levels in mice peripheral blood was assessed via flow cytometry 3–5 months after transplantation of human CD34+ stem cells. The gating strategy is presented in Figure 2A–E. Figure 3A and Figure 3B illustrate the variability between 10 and 16 individual mice receiving stem cells from two different donors. Transplantation of 75,000 hCD34+ cells yielded 20%–50% human CD45+ in the peripheral blood. All mice developed human B and T cells, including both CD4- and CD8+ T cells.
For atraumatic intravaginal exposures, the setup depicted in Figure 4 was used. Mice were anaesthetized in a closed chamber and kept under anesthesia during the exposure. Mice were held with the vagina facing up for 5 min after exposure to ensure virus solution engagement with mucosal surfaces.
Figure 5A shows the 64% HIV transmission success rate observed using this model. Mice were challenged with 21,400 infectious units (IU) of RHPA4259 intravaginally. This dose resulted in 64% of mice becoming HIV infected following vaginal exposure. For comparison, data from two different cohorts of mice exposed through an intravenous route are included. As expected, 100% of the mice became HIV+ with similar doses of RHPA and an additional strain (YU2) using this route.
Figure 5B depicts representative results from three mice that were infected with HIV and switched to a diet containing standard cART. Mice were switched back to regular mouse chow after 40 days of cART. In this assay setup, the limit for viral load detection was 725 copies/mL. Viral loads were all below the detection limit after 4 weeks of cART. After cessation of cART, the virus rebounded, mirroring clinical data22. Mice on cART tolerated the change in diet well as indicated in Figure 5C.
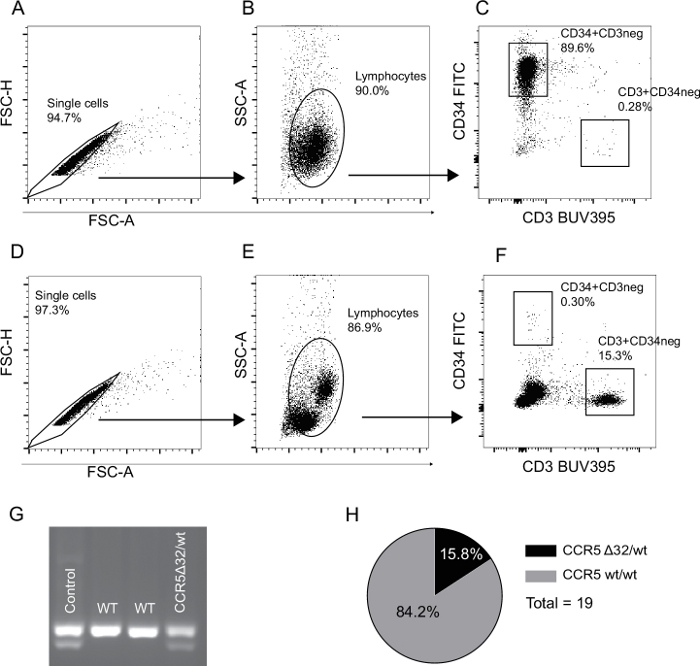
Figure 1: Representative flow cytometry gating strategy for validation of stem cell purity and CCR5 donor variant status. (A–C) The gating strategy used for the isolated CD34+ cell population. Doublets and debris are excluded in panel A and B respectively (FSC-A vs. FSC-H and FSC-A vs. SSC-A). (C) The frequency of CD34+ stem cells and CD3+ T cell contamination. (D–F) The CD34- flow-through gating strategy. Percentages in gates are calculated as a fraction of the parent population. (G) The results of a CCR5Δ32/wt PCR analysis. Lane 1: DNA from a human CCR5Δ32/wt donor, lanes 2 and 3: two CCR5wt/wt human stem cell donors, lane 4: A CCR5Δ32/wt human stem cell donor. (H) Frequency of the genotype CCR5Δ32/wt in our group of 19 stem cell samples is 15.8%. Please click here to view a larger version of this figure.
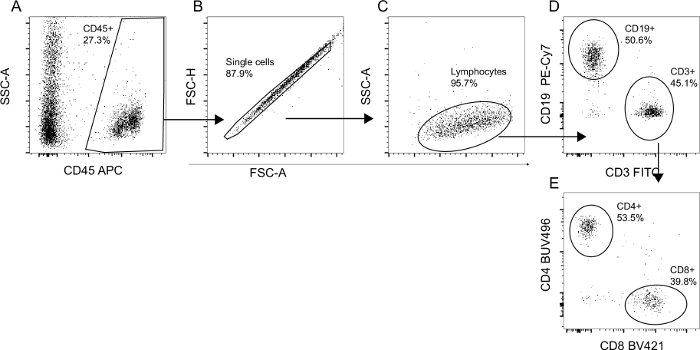
Figure 2: Flow cytometry gating strategy for validation of human cell engraftment and differentiation. The total mononuclear cell population from humanized mice was analyzed via flow cytometry. (A) The percentage of human CD45+ cells was determined as a fraction of the total recorded events. (B) Doublets were subsequently excluded based on FSC-A/FSC-H gating. (C) The true lymphocyte population was defined based on size and granularity. (D) Lymphocytes were then characterized as either CD3+ (T cells) or CD19+ (B cells). (E) CD3+ T cells were either CD4+ T cells or CD8+ T cells. Percentages in gates were calculated as a fraction of the parent population. Please click here to view a larger version of this figure.
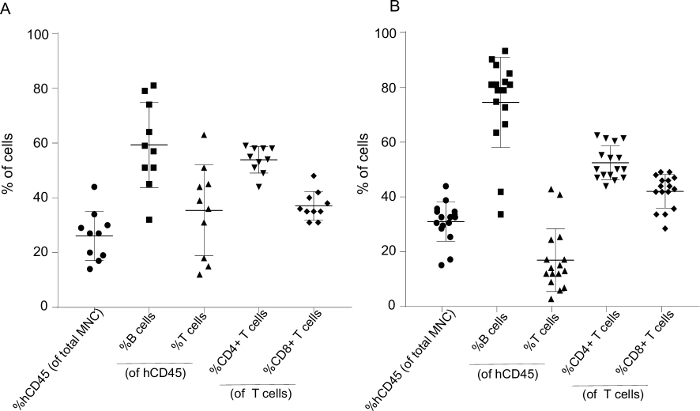
Figure 3: Representative humanization levels 4–5 months after stem cell transplantation with cell subtype fractions for 10 and 16 mice generated from two different human donors. (A) The mononuclear cell population (MNC) from 10 and (B) 16 humanized mice were analyzed via flow cytometry and gated as presented in Figure 2. The fraction of human CD45+ cells is presented as %hCD45 (of total MNC), and %B and %T cells as a fraction of hCD45. T cells were subsequently divided into %CD4 and %CD8. Each data point represents one mouse. Data is presented as mean ± SD. Please click here to view a larger version of this figure.

Figure 4: Experimental lab bench setup for intravaginal exposure of mice. Experimental setup for HIV exposure of humanized mice through the intravaginal route. The procedure is performed in a flow bench where all reagents and surfaces have been sterilized prior to use. Please click here to view a larger version of this figure.
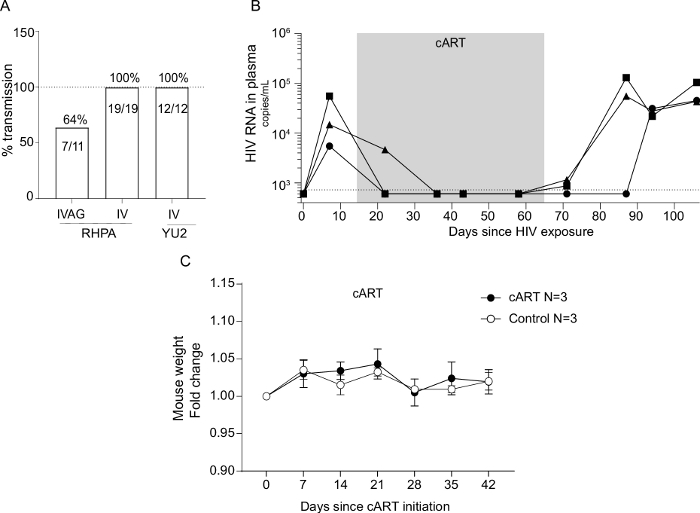
Figure 5: Rate of HIV strain transmission through different exposure routes and efficacy and safety of cART-containing chow in viral suppression. (A) Humanized NOG mice were successfully infected with two different strains of HIV through either the intravaginal or the intravenous route. Mice were exposed with 21,400 IUs of RHPA4259 intravaginally, 5,157 IUs IV with RHPA4259, or 3,000 IUs IV with YU2. Details regarding IV exposure of humanized mice are not included in this protocol. HIV infections were successfully treated with a cART regimen delivered through mouse chow. (B) The viral load decreased to below detection for all three mice on cART and rebound emerged after the cessation of cART. The dotted line indicates limit of quantification at 725 copies/mL. Mice fed with cART chow had similar weight development as mice housed on non-cART chow during the same time period, indicating no taste-preference or side effects of the cART diet. (C) Weights are presented as fold change compared to the start of cART. Each data point represents the mean of three animals ± SD. Please click here to view a larger version of this figure.
| Antibody target | Clone | Fluorophore |
| CD3 | clone SK7 | BUV395 |
| CD34 | clone AC136 | FITC |
| CD45 | clone 2D1 | APC |
Table 1: Antibodies used for determination of stem cell purity. Suggested multicolor flow cytometry panel for evaluation of stem cell purity. Listed are the antibody target, the clone, and the fluorophore.
| CCR5Δ32 detection | Primers |
| Forward primer | 5'CTTCATTACACCTGCAGCT'3 |
| Reverse primer | 5'TGAAGATAAGCCTCACAGCC'3 |
Table 2: CCR5Δ32 variant detection PCR primers. Forward and reverse primers used for detection of the 32 bp deletion in the CCR5 gene.
| No. of Cycles | 1 | 45 | 1 | ∞ |
| Temperature (°C) | 98 | 98/63/72 | 72 | 10 |
| Time | 30 s | 10 s/30 s/15 s | 5 min | ∞ |
Table 3: CCR5Δ32 variant detection PCR program. PCR cycling program used for amplification of the CCR5 gene.
| Antibody target | Clone | Fluorophore |
| CD4 | SK3 | BUV 496 |
| CD8 | RPA-T8 | BV421 |
| CD3 | OKT3 | FITC |
| CD19 | sj25c1 | PE-Cy7 |
| CD45 | 2D1 | APC |
Table 4: Antibodies used for determination of mouse humanization. Suggested multicolor flow cytometry panel for humanization. Listed are the antibody target, the clone, and the fluorophore.
| No. of Cycles | 1 | 1 | ∞ |
| Temperature (°C) | 51 | 80 | 4 |
| Time | 45 min | 15 min | ∞ |
Table 5: cDNA amplification program. Program used for amplification of complementary strand DNA to the viral RNA.
| HIV quantification | Primers |
| Forward primer | 5'AGGGCAGCATAGAGCAAAAA'3 |
| Reverse primer | 5'CAAAGGAATGGGGGTTCTTT'3 |
| FAM probe | 5'ATCCCCACTTCAACAGATGC'3 |
Table 6: HIV ddPCR primers. Primers and probes used for ddPCR amplification of viral cDNA.
| No. of Cycles | 1 | 39 | 1 | ∞ |
| Temperature (°C) | 95 | 95/54.5 | 98 | 4 |
| Time | 10 min | 30 s/1 min | 10 min | ∞ |
Table 7: HIV ddPCR program. PCR cycling program used for amplification of viral RNA.
| Raltegravir (RAL) | 4800 mg/kg |
| Tenofovir disoproxil fumarate (TDF) | 720 mg/kg |
| Emtricitabine (FTC) | 520 mg/kg |
Table 8: Mouse cART chow diet. Mouse chow diet was formulated as previously published21. The chow diet was made on a base of standard mouse chow, and after production, the food was γ-irradiated with 25 kGy and double-bagged. The chow was stored at -20 °C until use.
