1. Preparation and use of biosafety cabinet
- Wipe down biosafety cabinet with 70% ethanol.
- Close the sash and turn on the ultraviolet light in the biosafety cabinet for 15 min.
- After 15 min, turn off the ultraviolet light in the biosafety cabinet. Lift the sash and turn on the blower for 15 min.
- Wipe all items to be used in the biosafety cabinet with 70% ethanol prior to putting the items into the sterilized cabinet.
- Clean gloves or bare hands with 70% ethanol before working in the biosafety cabinet.
- Work in the center of the biosafety cabinet, away from the blower.
- Use unopened bottles of autoclaved sterile 10 mM MgCl2 and ultrapure H2O for experiments. Put bottles in the biosafety cabinet and only open them in the sterilized biosafety cabinet, not on the benchtop.
- Use dedicated glass pipettes and pipette tips for work in the sterilized biosafety cabinet. Ensure that these are only opened in the biosafety cabinet, never on the benchtop.
- After use of the biosafety cabinet, autoclave all waste (except bleach waste) and wipe down the surface with 70% ethanol.
2. Preparation of plant media
- Weigh out and dissolve 0.5x MS basal salts in ultrapure H2O. Weigh out 0.8% bacto agar and then add to dissolved 0.5x MS.
- Autoclave and allow the media to cool in 50 ˚C water bath for 1 h prior to pouring or pipetting.
- To ensure that plates are not overfilled, mark polystyrene disposable sterile 100 x 25 mm plates to a fill level of 40 mL. Pour media into 100 x 25 mm sterile plates in a sterilized biosafety cabinet.
3. Preparation of plant materials and growth conditions
- Place tomato seeds in a 2.2 mL microcentrifuge tube and add 2.0 mL of 50% bleach solution.
- Rock the tube on a rocker for 25 min.
- After 25 min, remove the seeds from the rocker and remove the bleach solution with a pipette in the sterile biosafety cabinet. Ensure that all the bleach is removed.
- Add 2 mL of sterile ultrapure H2O to wash the seeds. Invert the tube 5x.
- Remove the liquid from the tube with a pipette.
- Repeat steps 3.3–3.5 to wash the seeds 4x more.
- Add 2 mL of sterile ultrapure H2O and pour the seeds into an empty sterile Petri dish.
- Flame forceps in ethanol and allow to cool prior to transferring and evenly spacing seeds on 100 x 25 mm plates containing 0.5x MS + 0.8% agar media.
- Transfer 5–7 seeds in a line across the middle of one plate and seal the edges of the plates with surgical tape (1.25 cm x 9.1 m).
- Stratify the sterilized seeds at 4 ˚C in the dark for at least 3 days to synchronize germination. Ensure that the plates are stacked flat and face up, so that the seeds do not shift on the plate.
- Vertically orient the plates so that the roots will grow down along the surface of the plate, with the line of seeds oriented horizontally, when transferring to the growth chamber.
NOTE: Set the growth chamber to 22 ˚C and provide 16 h of light at a light intensity of ~200–220 µE meter-2 s-1 and 8 h of darkness. - Prior to flooding, grow seedlings for 10 days in the growth chamber at which point seedlings typically display fully emerged and expanded cotyledons and emerging first true leaves (Figure 1).

Figure 1: Developmental stage of typical 10-day-old tomato seedlings. Rio Grande-PtoR tomato seeds were sterilized, plated, and stratified for at least 3 days in the dark at 4 ˚C. The seedlings were grown on 0.5x MS plates for 10 days at 22 ˚C before being flooded. Typically, at 10 days the cotyledons are fully expanded, and the first true leaves are beginning to emerge. Please click here to view a larger version of this figure.
4. Preparation of King's B23 (KB) media
- Fill beaker with 500 mL of ultrapure H2O and stir on a stir plate.
- Completely dissolve 20 g of bacto peptone, 1.5 g of anhydrous K2HPO4, and 12.5 mL of glycerol in a beaker with ultrapure H2O.
- Pour the dissolved mixture into a 1 L graduated cylinder and bring up to a 1 L final volume with ultrapure H2O.
- Pour the broth back into the beaker and stir until mixed.
- Weigh out 7.5 g of bacto agar into two 500 mL glass bottles and add 500 mL of KB broth from step 4.4 into each bottle. Autoclave for 20 min.
- Remove the bottles from the autoclave and swirl gently to distribute the agar.
- Transfer the bottles to a 50 ˚C water bath for 1 h.
- After 1 h, transfer the bottle to the biosafety cabinet and under aseptic conditions, add 1,600 µL of sterile 1 M MgSO4, and appropriate antibiotics to the media.
NOTE: For rifampicin resistant strains PstDC3000 and Pst19, use rifampicin dissolved in dimethylformamide at a final concentration of 50 µg/mL. Use cycloheximide dissolved in ethanol at a final concentration of 50 µg/mL to prevent fungal growth on the plates. - Swirl the media gently to mix and then pour to cover the bottom of the plates.
- Allow at least 1 h for the plates to solidify before storing them upside down at 4 ˚C.
5. Maintenance of bacterial strains and culture conditions
- Maintain a glycerol stock from single colony of bacteria as 1 mL of saturated bacterial culture and 333 µL of sterile 80% glycerol at -80 ˚C.
- Patch bacteria (i.e., Pst19) from a glycerol stock onto KB agar with appropriate antibiotics (section 4).
- Allow the bacteria to recover for 2 days at 28 ˚C before streaking fresh bacteria onto selective KB agar using a flat, sterile toothpick.
- Streak fresh bacteria from the glycerol stock onto appropriate selective KB agar using a flat, sterile toothpick.
NOTE: Ensure that the patched glycerol stock is not more than 2 weeks old. - For PstDC3000, incubate the KB plate at 28 ˚C for 24 h prior to using bacteria in the flood experiment.
- For Pst19, incubate the KB plate at 28 ˚C for 48 h prior to using bacteria in the flood experiment.
6. Preparation of Pst19 inoculum
- Aseptically resuspend the bacteria in sterile 10 mM MgCl2 to an optical density at 600 nm (OD600) of 0.1, or approximately 5 x 107 colony forming units (CFU)/mL).
- Perform serial dilutions using sterile 10 mM MgCl2 solution in the biosafety cabinet. For Pst19, use a spectrophotometer to make inoculum with a starting concentration of OD600 = 0.1.
- For Pst19, make a 1/10 dilution from the initial resuspension at OD600 = 0.1 to obtain a serial dilution at a concentration of OD600 = 0.01.
- Using the serial dilution at OD600 = 0.01 from step 6.3, make a 3/4 dilution to obtain a final OD600 = 0.0075.
- Make a 1/10 dilution of non-ionic organosilicone surfactant copolymer C13H34O4Si3 (i.e., surfactant) in 10 mM MgCl2 and vortex for 15 s. Add the 1/10 stock of surfactant to the last serial dilution (OD600 = 0.0075) to a final concentration of 0.015% and swirl well to mix.
7. Preparation of PstDC3000 inoculum
- Aseptically resuspend bacteria in sterile 10 mM MgCl2 to an optical density at 600 nm (OD600) of 0.1 (approximately 5 x 107 CFU/mL).
- Perform serial dilutions using sterile 10 mM MgCl2 solution in the biosafety cabinet. For PstDC3000, use a spectrophotometer to make inoculum with a starting concentration of OD600 = 0.1.
- For PstDC3000, make a 1/10 dilution from the initial resuspension at OD600 = 0.1 to obtain a serial dilution at a concentration of OD600 = 0.01.
- Using the serial dilution at OD600 = 0.01 from step 3, make a 1/2 dilution to obtain a final OD600 = 0.005.
- Make a 1/10 dilution of surfactant in 10 mM MgCl2 and vortex for 15 s. Add the 1/10 stock of surfactant to the last serial dilution (OD600 = 0.005) to a final concentration of 0.015% and swirl well to mix.
8. Tomato seedling flood method
- Take the plates with the 10-day-old seedlings out of the growth chamber and put in the biosafety cabinet to prepare the plates for flooding.
- Remove the surgical tape from two plates.
- Set a timer for 3 min. Measure 6 mL of final inoculum (Pst19 OD600 = 0.0075 [section 6] or PstDC3000 OD600 = 0.005 [section 7]) and transfer 6 mL of inoculum to each plate with the 10-day-old seedlings.
- Gently push the seedlings down into the inoculum with a sterile pipette tip. Start the timer.
- Hold one plate in each hand. Tilt the front of the plate down to accumulate inoculum and mainly submerge the cotyledons and leaves of the seedlings.
- Swish side to side 5–7x and then tip the plates back to cover the roots and the whole plate.
- Tilt the plates down again to submerge the cotyledons and leaves, and repeat for a total of 3 min.
- Pour the inoculum off the plates, set the plates down on a flat surface and then pour off any residual inoculum a second time.
- Rewrap the plates with surgical tape and repeat steps 8.2–8.8 for any remaining plates.
- Re-incubate the plates in the growth chamber (see step 3.11 NOTE) after all plates have been flooded.
- Phenotype after 7–10 days for PstDC3000 or 10–14 days for Pst19 (section 11). If carrying out bacterial growth assays, collect leaf tissue after 4 days (sections 9 and 10) and then phenotype (section 11). Alternatively, perform phenotypic analysis and bacterial growth assays on separate sets of plants.
9. Surface sterilization of cotyledons for bacterial growth assay
- Four days after flooding and re-incubating the seedlings in the growth chamber (section 8), remove the plates with the tomato seedlings from the growth chamber.
- Number the individual seedlings on the bottom exterior of the plate where the seedling attaches to the plate for each genotype.
- Label sterile 1.5 mL microcentrifuge tubes with the individual seedling numbers and use clean forceps to drop one 3 mm sterile borosilicate bead into each tube for use with a bead beater. (See NOTE in step 10.1.)
- Pipette 200 µL of 10 mM MgCl2 into each tube and close tubes.
- Prepare 70% ethanol and pour 100 mL into a clean beaker. Pour 100 mL of sterile ultrapure H2O into a separate, clean beaker.
- Clean stainless steel straight fine-point forceps with serrated tips with ethanol. Open the plate slightly to allow aseptic removal of one cotyledon with the clean forceps.
- Pinch the petiole at the base of the cotyledon to remove a leaf and drop into the beaker with 70% ethanol to surface-sterilize for 10 s. Rinse the cotyledon in ultrapure H2O for 10 s.
- Place the cotyledon on a paper towel and blot dry with delicate science wipes.
- Individually weigh each cotyledon after surface sterilization and blotting, and record the weight.
- Place the cotyledon in a previously prepared 1.5 mL microcentrifuge tube (from steps 9.3 and 9.4) labelled with the corresponding genotype and individual number.
- Reseal the plates with sterile tape and re-incubate the seedlings in the growth chamber (see step 3.11 NOTE).
10. Bacterial growth assay
- Using samples from step 9.10, homogenize the tissue using the bead beater in 10 mM MgCl2 for 1–2 min. If the tissue is not adequately macerated, homogenize again.
NOTE: Many manufacturers produce bead beater homogenizers. The number and type of beads, as well as the homogenization time and speed (if programmable) should be optimized for each type of homogenizer. Ensure that the samples do not overheat during homogenization. - Add 800 µL of 10 mM MgCl2 to each tube containing macerated tissue from step 10.1 and invert several times to mix.
- Prepare serial dilutions for each sample in 10 mM MgCl2 in 96 well plate (100, 10-1, 10-2, 10-3, 10-4, 10-5) using a multichannel pipette (Figure 2A).
- Pipette 5 µL from each dilution series using a multichannel pipette onto a KB agar plate (150 mm x 15 mm) with cycloheximide and appropriate selection for the bacterial strain of interest (see step 4.8 NOTE). Let the plates dry completely.
- Incubate the plate upside down at 28 ˚C for 36 h, then visualize (Figure 2B) the colonies on the plates using a dissecting microscope to determine if the colonies are large enough to count.
NOTE: If the colonies are not large enough, re-incubate the plates and recheck the size of the colonies every few hours. Typically, the colonies are countable ~36–48 h after incubation.
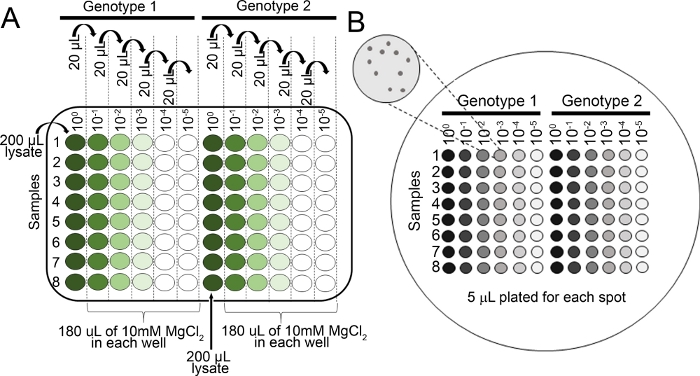
Figure 2: Serial dilutions for seedling bacterial growth assays. (A) Macerated leaf tissue from infected plants is diluted prior to colony counting. Dilutions are performed in a 96 well plate (100 is undiluted). Typically, dilutions are made from 10-1 to 10-5. (B) Plating dilutions for bacterial colony counts. A total of 5 µL of each column of the dilution series is plated, from most dilute to most concentrated. After the colonies have fully dried, the plate is incubated at 28 ˚C for 36–48 h. Colonies are counted under a 10x dissecting microscope. Please click here to view a larger version of this figure.
- Count the colonies under a dissecting microscope before they merge (Figure 2B). Count the colonies from the dilution series plates with fewer than 100 colonies.
- After obtaining colony counts (Figure 2B), normalize the counts to 0.1 g of tissue for seedlings and convert to log bacterial growth (Table 1).
NOTE: The average mass of one Moneymaker-PtoS cotyledon is 0.1 g and is empirically determined22.
| Genotype1 Column A | Tissue Weight (g) Column B | # of Colonies in a spot Column C | Dilution factor for spot2 Column D | Adjusted # of Colonies3 Column E | Dilution factor for serial dilution Column F | Total # of Colonies Column G (cfu/0.01 g)4 | Average # of Colonies (cfu/0.01 g) Column H | Average Log Growth (cfu/0.1 g (log10)) Column I |
| Sample 1 | 0.04 g | 10 | 200 | calculated as: (C2 x 0.1 g) / B2 = 25 | 1000 | calculated as: (D2 x E2 x F2) = 5000000 | average for sample 1 through last sample: (ie. average G1:G3) = 7000000 | log of average ie. log(H2) = 6.85 |
| Sample 2 | 0.03 g | 15 | 200 | 50 | 1000 | 10000000 | ||
| Sample 3 | 0.02 g | 6 | 200 | 30 | 1000 | 6000000 | ||
| 1Data shown for 3 samples | ||||||||
| 2Based on plating 5 µL x 200 for 1 mL | ||||||||
| 3Cotyledons are too small to core so colony counts were normalized to 0.1 g of tissue based on the average mass of one MoneyMaker-PtoS cotyledon (data not shown) | ||||||||
| 4Adjusted per mL based on volume plated | ||||||||
Table 1: Sample calculations for seedling bacterial growth assay. Sample calculations demonstrate how to normalize bacterial counts and determine log bacterial growth.
- For wild accessions and other lines with complex genetic backgrounds, correlate the level of bacterial growth in individual seedlings with their phenotype as described in section 11.
11. Phenotyping for resistance
- Remove the plates from the growth chamber and phenotype individual seedlings for death (due to disease) or survival (due to resistance) after 7–14 days.
- Phenotype plants infected with a highly virulent strain such as PstDC3000 earlier, at 7–10 days after flood inoculation.
- Phenotype plants infected with Pst19 at 10–14 days after flood inoculation.
- Determine a scoring system based on the range of resistance phenotypes observed. Record binary phenotypes for cultivars, isogenic lines, and wild accessions with consistent, strong to intermediate resistance phenotypes (Figure 4A, 4B).
- If the seedling displays new growth from the apical meristem within the time frame for phenotyping, count it as a survival. If the seedling has a brown apical meristem and displays no new, green vegetative growth, count it as a death (Figure 3).
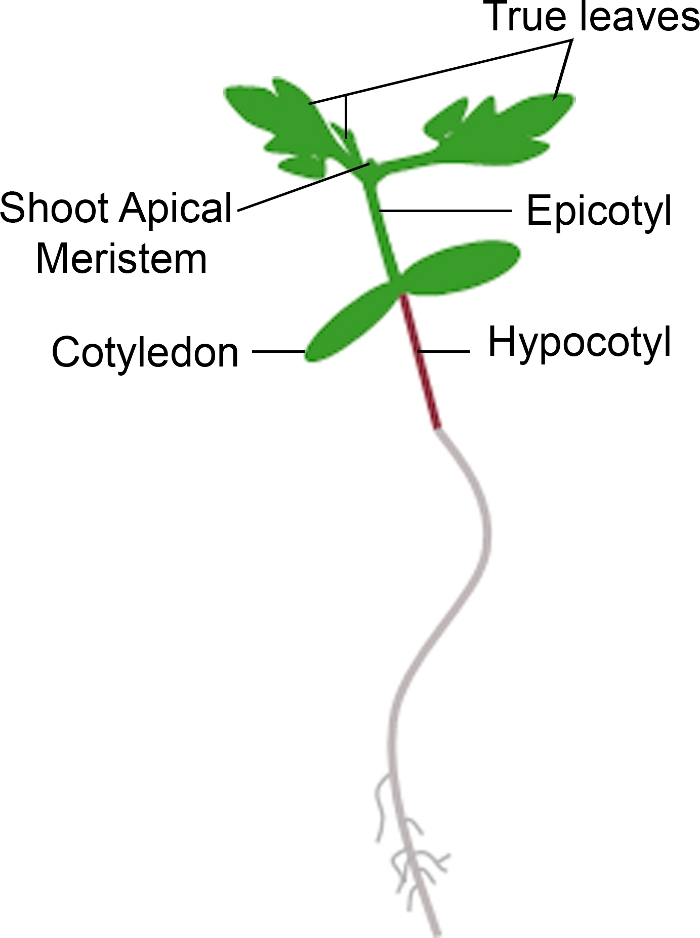
Figure 3: Schematic representation of a tomato seedling. Different parts of a tomato seedling are depicted, including the hypocotyl, cotyledons, epicotyl, shoot apical meristem, and true leaves. Please click here to view a larger version of this figure.
- Record phenotypes on a disease spectrum for populations, such as F2 mapping populations, with a wide range of resistance phenotypes (Figure 4C).
- Carefully monitor the seedlings for the appearance of disease symptoms and death to identify the appropriate window for phenotyping.
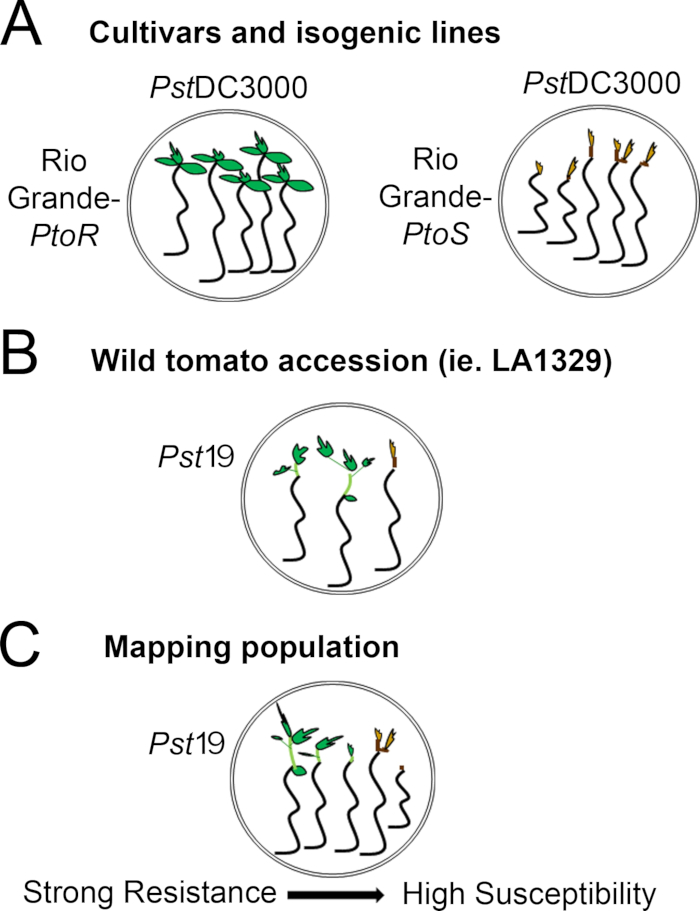
Figure 4: Schematic representation of expected phenotypes for seedling resistance and death in various genetic backgrounds. (A) Seedlings of Rio Grande-PtoR and the near-isogenic cultivar Rio Grande-PtoS are displayed 7 days after flooding with PstDC3000 (OD600 = 0.005) + 0.015% surfactant. Rio Grande-PtoR displays consistent resistance, and Rio Grande-PtoS displays consistent susceptibility to infection with PstDC3000. These lines give rise to discrete and binary phenotypes. (B) Seedlings of a wild accession, such as Solanum neorickii LA1329, are shown 10 days after flooding with Pst19 (OD600 = 0.0075) + 0.015% surfactant. Seedlings display phenotypic variability but were recorded as binary phenotypes. The amount of phenotypic variability and the method of phenotyping (binary resistance or resistance spectrum) will depend on the particular accession tested. (C) Mapping populations generated by outcrossing wild accessions to susceptible cultivars may display a wider spectrum of phenotypes in F2 segregating populations. In this case, it may be most appropriate to record seedling phenotypes on a spectrum. Highly susceptible seedlings from a mapping population may be phenotyped for death as early as day 7 when flooded with Pst19, and typically show a brown apical meristem, no to very little extension of the epicotyl, and no new, green vegetative growth. The apical meristem of susceptible seedlings may stay green or very light brown for more time, and there may be some extension of the epicotyl and very little vegetative growth, which turns brown and arrests by day 10. Individual seedlings can be phenotyped for resistance based on the amount of new and ongoing vegetative growth by day 14. Seedlings can then be grouped based on the phenotypes described above into different categories of resistance such as weak, medium, or strong resistance. Please click here to view a larger version of this figure.
Detection of PtoR-mediated immunity in cultivars and isogenic lines using the seedling resistance assay
Figure 5 shows representative results for Moneymaker-PtoR and Moneymaker-PtoS cultivars 7–10 days after flooding with PstDC3000. Prior to infection, 10-day-old seedlings displayed fully emerged and expanded cotyledons and emerging first true leaves. The seedlings were flooded with 10 mM MgCl2 + 0.015% surfactant as a negative control (data not shown) and PstDC3000 at an optical density of 0.005 + 0.015% surfactant. The seedlings were phenotyped 7–10 days after flooding (Figure 5). Individual seedlings from genotypically homogenous lines, such as Moneymaker-PtoR and Moneymaker-PtoS give highly consistent and binary phenotypes in the seedling flood assay. When Moneymaker-PtoR, which carries the Pto/Prf gene cluster (n = 5), was treated with PstDC3000 at the optimal concentration of OD600 = 0.005, resistance due to PtoR-mediated immunity was strong and was typified by new, green vegetative growth in all individuals22. Near-isogenic Moneymaker-PtoS seedlings (n = 5), which cannot recognize the PstDC3000 effectors AvrPto or AvrPtoB, died quickly within 7 days after flooding and characteristically had brown apical meristems, bacterial speck, chlorosis, and no signs of new, green vegetative growth (Figure 5).
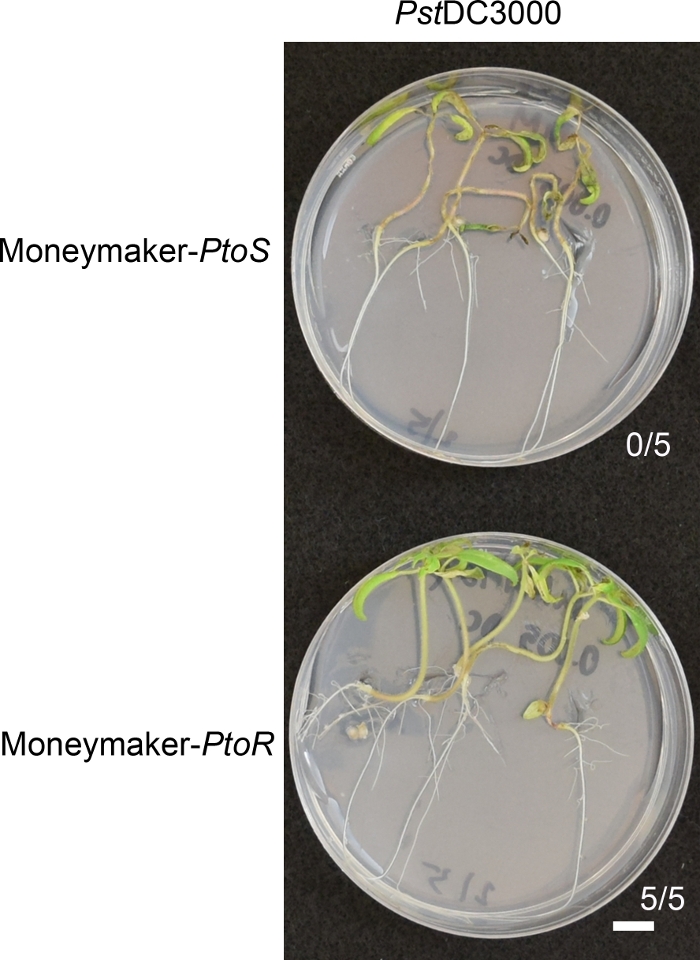
Figure 5: Phenotypic characterization of resistance or disease symptoms 7–10 days post-infection in a cultivar. Moneymaker-PtoR and Moneymaker-PtoS tomato seedlings were grown on 0.5x MS plates for 10 days before being flooded with P. syringae pv. Tomato DC3000 (OD600 = 0.005) + 0.015% surfactant. Moneymaker-PtoR seedlings survived (n = 5) and Moneymaker-PtoS seedlings (n = 5) died. The number of surviving seedlings for each genotype out of the total number tested is shown. Scale bar = 1 cm. Please click here to view a larger version of this figure.
Phenotypic screening of wild accessions using the seedling resistance assay
Figure 6 shows representative results for seedlings of susceptible and resistant accessions 10–14 days after flooding with Pst19. Susceptible accessions include RG-PtoR, S. pimpinellifolium LA1375, and S. pimpinellifolium LA1606, and resistant accessions include S. neorickii LA1329. Ten-day-old seedlings were flooded with 10 mM MgCl2 + 0.015% surfactant as a negative control, and Pst19 at an optical density of 0.0075 + 0.015% surfactant. The seedlings were phenotyped at least 10 days after flooding, as Pst19-infected seedlings died more slowly than PstDC3000-infected seedlings. Mock-inoculated seedlings were green, healthy, and actively growing. This control is important to ensure that the accessions are not sensitive to the concentration of surfactant, and to ensure there is no bacterial contamination. Susceptible accessions (Rio Grande-PtoR [n = 7], S. pimpinellifolium LA1375 [n = 7], and S. pimpinellifolium LA1606 [n = 5]) were dead, had brown apical meristems, and lacked new growth 10–14 days after inoculation with Pst19. In contrast, two S. neorickii LA1329 (n = 3) seedlings displayed a high level of new, green growth and survived infection with Pst19 (Figure 6). Three LA1329 seedlings did not germinate. Typically, 5–7 individuals were screened for each accession in a primary screen to determine the prevalence of resistance in the population. When a more genetically complex wild accession, such as LA1329, is flooded with Pst19, the resistance phenotypes display slightly more variability among individual seedlings, compared to Moneymaker-PtoR treated with PstDC3000. However, the resistance phenotypes were usually less variable than those seen in F2 mapping populations. Thus, binary phenotyping criteria was used for LA1329.
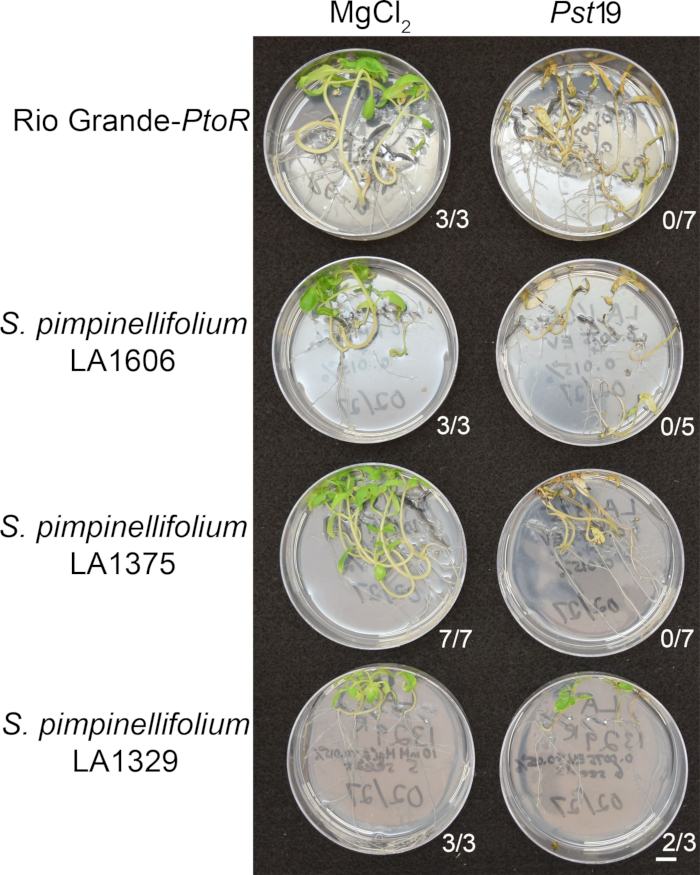
Figure 6: Phenotypic characterization of resistance or disease symptoms 10–14 days post-infection in wild accessions. Rio Grande-PtoR, S. pimpinellifolium LA1606, S. pimpinellifolium LA1375 and S. neorickii LA1329 tomato seedlings were grown on 0.5x MS plates for 10 days, and then flooded with Pst19 (OD600 = 0.0075) + 0.015% surfactant. The number of surviving seedlings for each wild accession out of the total number tested is shown. Scale bar = 1 cm. Please click here to view a larger version of this figure.
Quantitative assessment of bacterial growth using the seedling flood assay
To confirm that the observed resistance in LA1329 to Pst19 resulted in lower bacterial growth, bacterial growth assays were carried out in tomato seedlings. The level of Pst19 growth in Moneymaker-PtoS and S. neorickii LA1329 was determined 4 days post-infection. Moneymaker-PtoS is a near-isogenic line with consistent susceptibility among individual seedlings. Wild accessions such as S. neorickii LA1329 are often more genetically complex. LA1329 displays approximately 60% resistance to Pst19 across the population22. Because seedlings may drop their cotyledons after infection, one seedling was grown on each plate to correlate bacterial growth in the harvested cotyledon with overall seedling survival or death as determined phenotypically at least 10 days after flooding. The bacterial counts on day 4 for each seedling were normalized to 0.01 g of tissue and converted to log growth (CFU/0.01 g(log10)). Log growth for phenotypically resistant LA1329 seedlings (LA1329RES) or phenotypically susceptible seedlings (LA1329SUS) were separately pooled and compared to each other and the susceptible cultivar Moneymaker-PtoS. For example, there was a 1.7 log difference in bacterial growth between LA1329RES (log 6.3) and LA1329SUS (log 8.0), and a 1.6 log difference between LA1329RES (log 6.3) and Moneymaker-PtoS (log 7.9) (Figure 7). Therefore, phenotypic resistance correlated with quantitative resistance in the seedling assays.
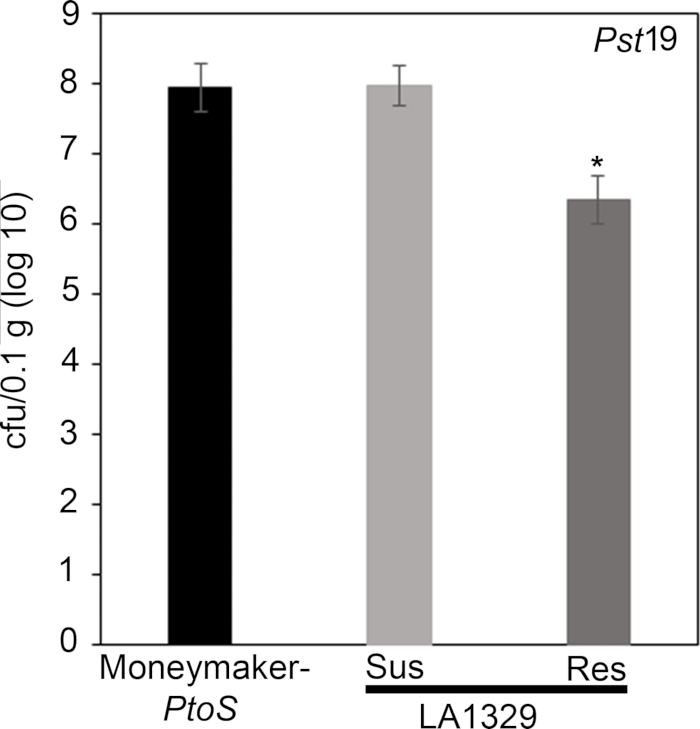
Figure 7: Resistant Solanum neorickii LA1329 seedlings support lower bacterial growth than Moneymaker-PtoS or susceptible S. neorickii LA1329. Bacterial counts were determined 4 days post-inoculation from S. neorickii LA1329 (n = 14) and Moneymaker-PtoS (n = 10) seedlings infected with Pst19 and normalization was performed to 0.1 g of tissue. For LA1329, the two phenotypic groups, susceptible (SUS) or resistant (RES), were observed and counted separately. Above the bar * = statistically significant difference determined by a one-factor analysis of variance. A general linear model procedure (p < 0.001) followed by a multiple comparison of means using Tukey's post hoc test was used. Error bars = standard error. The figure indicates one representative experiment. Please click here to view a larger version of this figure.
