Immunoblotting with antibodies against the protein of interest reveals the palmitoylated state (non, singly, doubly, etc.) in mouse brain lysates as determined by mobility shift compared with samples in which HAM was not included. Previously, we had demonstrated that SynDIG1 was palmitoylated at two sites using the ABE assay6; however, we could not determine whether both sites were modified in brain tissue. Here we show that SynDIG1, SynDIG4/Prrt1, and Prrt2 are palmitoylated in mouse brain and that they have two potential palmitoylation sites (Figure 1). Interestingly, each protein has a different palmitoylation state in mouse brain based on the signal for the non, singly and doubly palmitoylated protein. Densitometry analysis of blots provides quantitative information about the palmitoylated state.
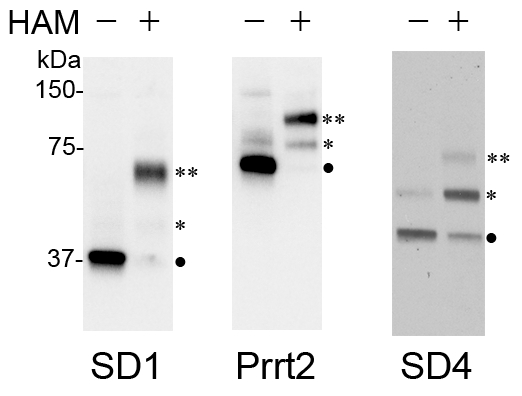
Figure 1: Detection of PEGylated protein species in mouse membrane preparations. Mouse brains were dissected rapidly and homogenized immediately. P2 membrane fractions were subjected to the APEGs assay as described in this protocol. In this experiment, postnatal day 18 (P18) were separated using a 10% SDS-PAGE gel and immunoblotted and probed with antibodies against SynDIG1 (SD1), Prrt2, and SynDIG4 (SD4). Symbols indicate protein that is not palmitoylated (•), palmitoylated singly (*), or doubly (**). The presence of faint bands in the HAM (-) conditions indicates either an incomplete reduction of disulfide bonds or incomplete blockage of free cysteines by NEM. Please click here to view a larger version of this figure.
| Buffer | Components | Working conc. | Comments |
| Homogenization | 0.32 M sucrose, 1 mM Tris pH 7.2, 1 mM MgCl2 | Add PMSF and protease inhibitors immediately before use. | |
| Buffer A | PBS pH 7.2, 5 mM EDTA, 4% SDS (w/v) | Add PMSF and protease inhibitors immediately before use. | |
| TCEP solution | 500 mM TCEP in ddH2O (pH 7.2) | 25 mM | Add 10 N NaOH to increase pH. |
| NEM solution | 2.0 M NEM in 100% ethanol | 50 mM | Prepare fresh. |
| Buffer H (HAM+) | 1.33 M HAM, pH 7.0, 5 mM EDTA, 0.2% Triton X-100 (w/v) | 1.0 M | Prepare fresh. |
| Buffer T (HAM-) | 1.33 M Tris-HCl, pH 7.0, 5 mM EDTA, 0.2% Triton X-100 (w/v) | ||
| mPEG-5k | 100 mM mPEG-5k in ddH2O | 20 mM | Mix well. Highly viscous. |
Table 1: Solutions used for APEGS assay. Homogenization buffer and buffer A can be prepared ahead of time; however, add protease inhibitors and phenylmethylsulfonyl fluoride (PMSF) immediately before use. All other solutions should be prepared fresh.
