Lymphatic vessel suture has been used in previous studies15,16,17,19, where it served as an important tool to study the function of lymph flow before the molecular biology of lymphatic vessels was better understood. Blocking lymph flow interrupts LN homeostasis, which leads to HEVs losing the critical gene expression needed for optimal lymphocyte homing to the LN15,16,17. Since then, it took another two decades to demonstrate that DCs traveling with lymph are crucial in maintaining the HEV gene expression profile and lymphocytes homing to the LN13. The shear stress provided by the lymph flow is critical to stimulate chemokine expression in the LN. Blocking lymph flow interrupts chemokine CCL21 expression in the LN19, which is critical in directing DC and T cell positioning in the LN. Therefore, interrupted flow may compromise DCs and T cells positioning in the LN8,18.
Immediately after the surgery, a small molecular weight fluorescent tracer, FITC, was used to track lymph flow. FITC (10 µL of 2% FITC) was injected intradermally in the footpad of the sham control and the lymphatic sutured leg. The draining pLNs were collected 2, 6, and 12 h later. The draining pLNs were embedded in OCT, and 20 µm frozen sections were prepared. Confocal images showed substantially reduced FITC accumulation in the pLNs after suture. The residual FITC in the pLNs was preferentially accumulated in the LN sinuses (Figure 2).
How the lymph flows through the adipose tissue surrounding the LN was investigated using lymphatic suture. The afferent lymphatic vessels leading to the pLNs were sutured to block lymph flow, and it was determined that the perinodal adipose tissue could support a small amount of lymph flow when lymphatic vessels were blocked21. The lymph flow through the adipose tissue to the capsule of the LN was mapped; it appeared to feed into the LN sinuses. Small amounts of lymph may have flowed into the LN sinuses over time (Figure 3).
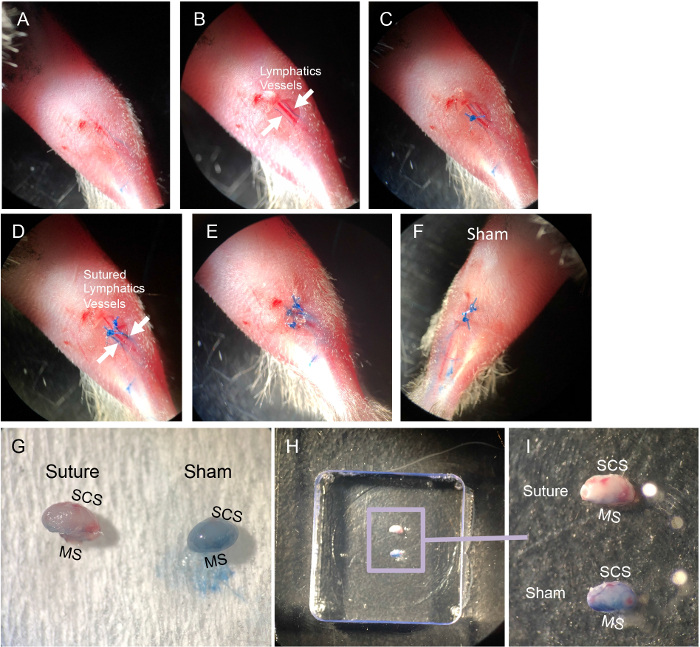
Figure 1: Steps of popliteal LN (pLN) afferent lymphatic vessel suture. Briefly, after mice were anesthetized with a ketamine and xylazine mixture, their legs were shaved, and the residual fur was removed by a depilatory cream. The right leg was used for suture and the left leg was the sham control. The right side of the footpad was intradermally injected with 5 µL of 1% Evans Blue dye prepared in PBS. (A) By gently massaging the footpad, Evans Blue dye filled the afferent lymphatic vessels. (B) A small skin cut was performed 5 mm away from the pLN to expose the lymphatic vessels, which are indicated by the two white arrows. (C,D) Both afferent lymphatic vessels were sutured. (E) The skin excision was closed by sutures. (F) The control leg received Evans blue injection, skin excision, and suture closure without suturing the lymphatic vessels. (G) The success of the lymph flow blockage was indicated by Evans blue dye, which entered the pLN of the control leg but not the sutured leg. (H,I) The collected pLNs were embedded in OCT compound with subcapsular sinus (SCS) and medullary sinus (MS) facing the side of the cryomold before snap freezing in liquid nitrogen. Please click here to view a larger version of this figure.
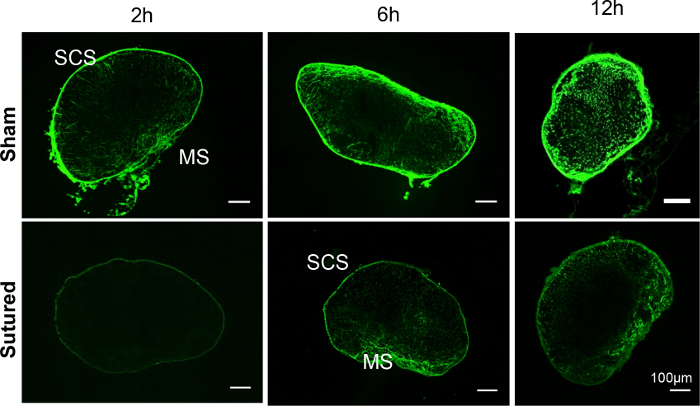
Figure 2: FITC distribution in the draining pLNs of the sham or sutured leg. Confocal images of pLNs collected 2, 6, and 12 h after FITC injection showed substantially reduced FITC accumulation in the pLNs after suture. The residual FITC in the pLNs was preferentially found in the LN sinuses. Please click here to view a larger version of this figure.
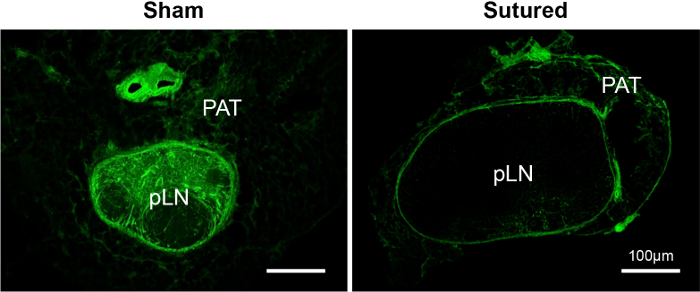
Figure 3: FITC distribution in the perinodal adipose tissue (PAT), around the draining pLNs of the sham or sutured leg. Confocal images of the PAT and the LN showed that FITC enters the PAT and the LN sinuses but was not effectively distributed throughout the LN when lymphatic vessels were blocked. Please click here to view a larger version of this figure.
