We constructed pSIN-NTCP-EGFP plasmid expressing NTCP and EGFP fusion and with puromycin resistance. The plasmid was transfected into HepG2 and 293T cells to construct stable cell lines HepG2-NE and 293T-NE expressing NTCP and EGFP. Plasmids (pSIN-HNF4α, pSIN-RXRα, pLV-PPARα-puro-flag) with puromycin resistance and expression were transfected into 293T-NE cells to construct a stable cell line expressing 4 host genes9. The expression of NTCP-EGFP can be observed by green fluorescence, and verified by qPCR and western blot (data not shown, but see previous work 9).
Primary antibody of HBcAg and DAPI incubated with fixed cells showed a distinct staining when observed using 10x objective on a fluorescent inverted microscope. Nuclear localization was confirmed by staining with DAPI (Figure 3 and Figure 4-the third column). NTCP-EGFP is expressed on the cell membrane and can be located by the green fluorescence (Figure 3 and Figure 4-the second column). The expression sites of HBcAg can be located by red fluorescence (Figure 3 and Figure 4-the first column). When DAPI, NTCP and HBcAg are in the same cell, it means that the cell has been successfully infected with HBV (Figure 3 and Figure 4-the last column).
The CsA group was the negative control. CsA prevented HBV from entering cells by blocking NTCP. As a positive control, infected HepG2-NTCP-EGFP cells can express HBcAg. HBcAg was detected in 293T-NE-3NRs cells but not in 293T-NE cells, indicating NTCP is not the only factor essential for HBV infection in 293T. The immunofluorescence of cells infected with HepG2.2.15 supernatant concentrate was the same as that of cells co-cultured with HepG2.2.15 cell. It showed that HBcAg was expressed in 293T-NE-3NRs and HepG2-NE cells, this suggests that the four host genes (NTCP, HNF4α, RXRα and PPARα) facilitates HBV infection. But signals for HBcAg in 293T-NE-3NRs were lower compared to HepG2-NE, indicating HBV infection may be also affected by other host factors.
| Quantitative reference | Quantitative reference | Quantitative reference | Quantitative reference | Negative control | HBV positive control | |
| 1 | 2 | 3 | 4 | |||
| Ct | 20.1527 | 17.6647 | 14.5816 | 12.0409 | none | 12.3865 |
| GEq/mL | 2*10^3 | 2*10^4 | 2*10^5 | 2*10^6 | —— | —— |
Table 1: Ct values for the Quantitative references.
| HepG 2.2.15 supernatant |
1/20 HepG 2.2.15 concentrate |
HepG 2.2.15 concentrate |
|
| Ct | 17.716 | 13.1556 | —— |
| GEq/mL | 1.65*10^4 | 7.53*10^5 | 1.5*10^7 |
Table 2: Ct values for HBV DNA obtained from HepG2.2.15 supernatant. Values represent Ct values for HBV DNA obtained from original supernatant, 1/20 dilution or its concentrate.
| 293T-NE | HepG2-NE | 293T-NE-3NRs | |||
| HBV | HBV | HBV+CsA | HBV | HBV+CsA | |
| HBV GEq/cell | 150 | 150 | 150 | 150 | 150 |
| HBV µL | 100 | 100 | 100 | 100 | 100 |
| CsA µL (5 µM) | 0 | 0 | 0.5 | 0 | 0.5 |
| DMSO µL | 10 | 10 | 9.5 | 10 | 9.5 |
| 40 % PEG8000 µL | 50 | 50 | 50 | 50 | 50 |
| Infection medium µL | 340 | 340 | 340 | 340 | 340 |
| Total µL | 500 | 500 | 500 | 500 | 500 |
Table 3: Infection complex. The total volume used was 500 µL. Cyclosporine A was used as a negative control.
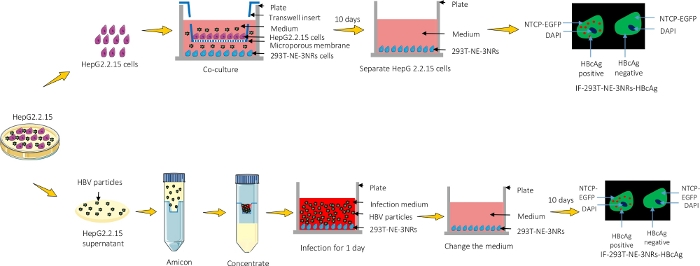
Figure 1: A graphical representation of the protocol. HepG2.2.15 were either co-cultured with 293T-NE-3NRs and infection was observed using immunofluorescence microscopy or the supernatant containing viral particles was concentrated and introduced into 293T-NE-3NRs cultured cells. Please click here to view a larger version of this figure.
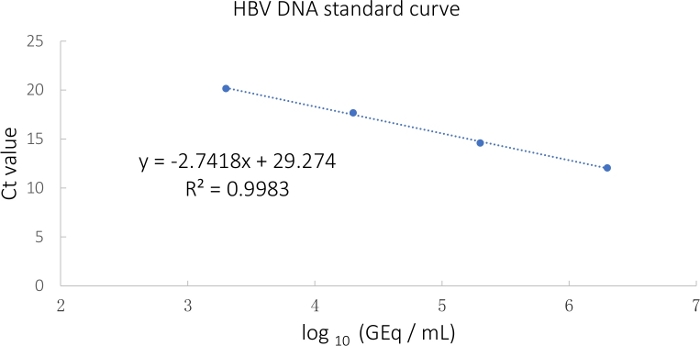
Figure 2: HBV standard curve calculated from the Ct values. Please click here to view a larger version of this figure.
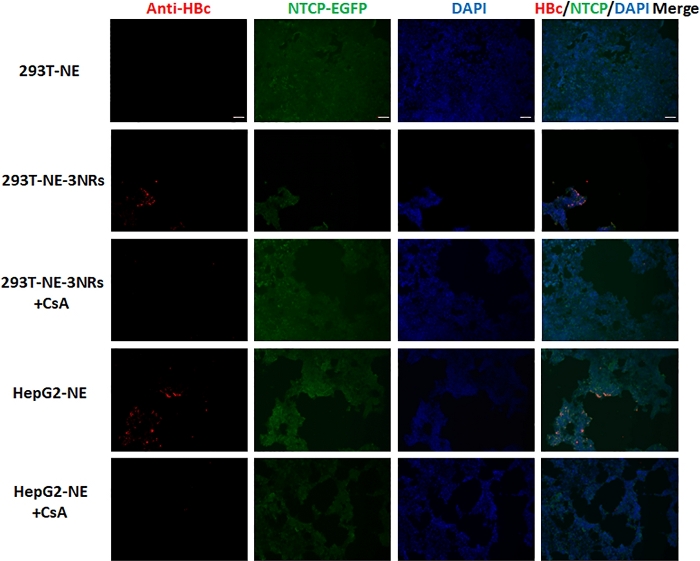
Figure 3: HepG2.2.15 supernatant concentrate was used to infect 293T-NE, 293T-NE-3NRs and HepG2-NE cells with 150 GEq per cell. HBcAg expression was identified by immunofluorescence assay. HBcAg was expressed in 293T-NE-3NRs and HepG2-NE cells, whereas, no expression was observed in these cells treated with CsA (HBV entry inhibitor) and 293T-NE cells. Scale bar = 100 µm. Please click here to view a larger version of this figure.
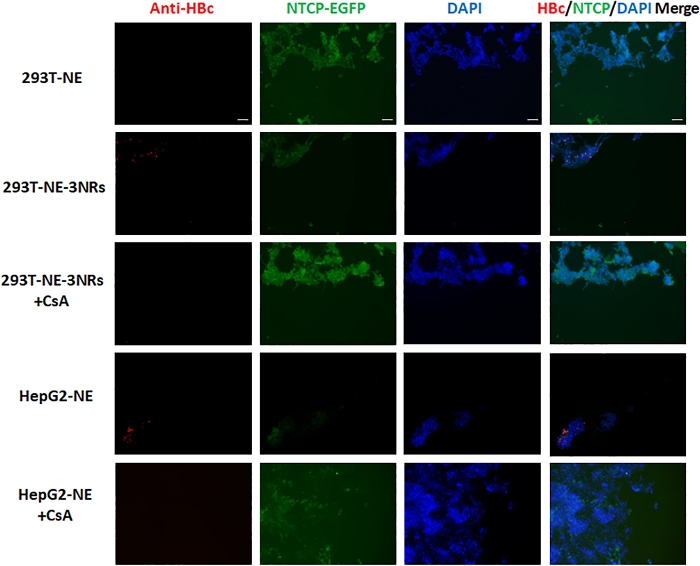
Figure 4: 293T-NE, 293T-NE-3NRs and HepG2-NE co-cultured with HepG2.2.15 cells. HBcAg expression was identified by immunofluorescence assay. HBcAg was expressed in 293T-NE-3NRs and HepG2-NE cells, whereas, no expression was observed in these cells treated with CsA (HBV entry inhibitor) and 293T-NE cells. Scale bar = 100 µm. Please click here to view a larger version of this figure.
