Tritonia
Skin contact with its seastar predator triggers Tritonia diomedea’s escape swim, consisting of a rhythmic series of whole-body flexions that propel it away to safety (Figure 3A). In isolated brain preparations, a brief stimulus to pedal nerve 3 (PdN3) elicits the rhythmic swim motor program (SMP) for this behavior, which is readily recognizable in optical recordings from the pedal ganglia. Figure 3B depicts the layout of a VSD imaging experiment designed to record the firing activity of neurons on the dorsal surface of the left Tritonia pedal ganglion, over which the PDA was positioned, as a stimulus to the contralateral (right) PdN3 elicits the SMP. Raw and filtered data (bandpass Butterworth filter, 5 and 100 Hz cutoffs) from 20 diodes recording activity before, during and after stimulation of PdN3 are shown in Figures 3C,D respectively. The nerve stimulus was delivered 20 s into the 2 min file. Immediately after acquisition, the signals measured by all 464 diodes of the recording array can be topographically displayed over an image of the preparation in the imaging software (Figure 3E). At this point, many traces contain spikes recorded redundantly from the same neurons, and some traces contain spikes from more than one neuron. Spike-sorting the filtered diode traces with ICA yielded 53 unique neuronal traces, 30 of which are shown in Figure 3F. The kinetics of individual spikes can be appreciated in Figure 3G, which expands an excerpt of four traces from Figure 3F (red box); the accuracy of the ICA spike-sorting algorithm was previously verified using simultaneous sharp electrode recordings, which showed that all spikes in the sorted traces correspond to intracellularly recorded spikes from individual neurons11,14.
Aplysia
A strongly aversive tail stimulus to Aplysia californica elicits a stereotyped two-part rhythmic escape response15. The first phase of the response is a gallop of several cycles of head lunges and tail pulls that move the animal quickly forward. This is typically followed by a period of crawling, involving repeated waves of head-to-tail muscular contractions that drive the animal forward at a slower speed for several minutes (Figure 4A). To capture these escape motor programs in optical recordings, the PDA was focused on the dorsal surface of the right pedal ganglion in an isolated brain preparation, and a suction electrode was placed on the contralateral (left) pedal nerve 9 (PdN9; Figure 4B). One minute into a continuous 20 min optical recording (Figure 4C), PdN9 was stimulated to elicit the gallop-crawl motor program sequence. The probabilistic Gaussian spatial distributions of the signals from all 81 recorded neurons were mapped onto the ganglion (Figure 4D). Dimensionality reduction applied to the full recording revealed that the gallop (cyan) and crawl (dark blue) phases of the escape program occupied distinct areas and formed different trajectories, spiral- and loop-like, respectively, in principal component space (Figure 4E).
Three videos based on the Aplysia recording depicted in Figure 4 demonstrate further types of analyses that can be performed on such data sets. Video 1 animates the firing of all recorded neurons over the full duration of the recording. The initial post-stimulatory period of the escape motor program was characterized by a gallop, in which activity in the ganglion was marked by alternating bursting of different functional clusters (Video 2). The gallop subsequently transitioned to a crawl, in which the activity across neuronal clusters remained broadly phasic but assumed a counterclockwise rotational trajectory in the ganglion (Video 3). The latter two videos also incorporate consensus clustering, which reveals the firing and locations of the different functional ensembles for the gallop and crawl phases of the escape response separately. Note that many neurons assigned to the same cluster in both the gallop and crawl phases exhibited physical proximity to one another in the ganglion, consistent with prior findings12.
Berghia
The aeolid nudibranch Berghia stephanieae (Figure 5A) represents a new model system for neuroscience. The imaging setup for a typical Berghia experiment is shown in Figure 5B. To elicit widescale neuronal activity, a suction electrode was placed on the most prominent left pedal nerve, and a nerve stimulus was delivered 30 s into a 2 min recording. ICA-processed traces revealed both spontaneous and stimulus-evoked activity in 55 neurons (Figure 5C). Community detection via consensus clustering identified ten distinct functional ensembles, which are depicted in Figure 5D in a network graph and in Figure 5E, which reorganizes the traces shown in Figure 5C based on their clustering assignments. Gaussian distributions of the signals from all recorded neurons are superimposed on an image of the preparation in Figure 5F to indicate the positions of all 55 recorded neurons.

Figure 1: Views of the optical imaging rig and photodiode array (PDA). (A) The optical imaging rig, featuring the PDA, digital camera, microscope, and stage. (B) The internal design of the PDA, in which fiber optics connect the imaging aperture to 464 photodiodes. A row of amplifiers is located above the photodiodes. (C) The hexagonal face of the imaging aperture, onto which the area being imaged is focused. Please click here to view a larger version of this figure.

Figure 2: A flowchart illustrating the essential workflow in obtaining optical recordings. The essential steps in the VSD imaging protocol, from dissection and staining through the details of imaging, are depicted in this flowchart. Please click here to view a larger version of this figure.
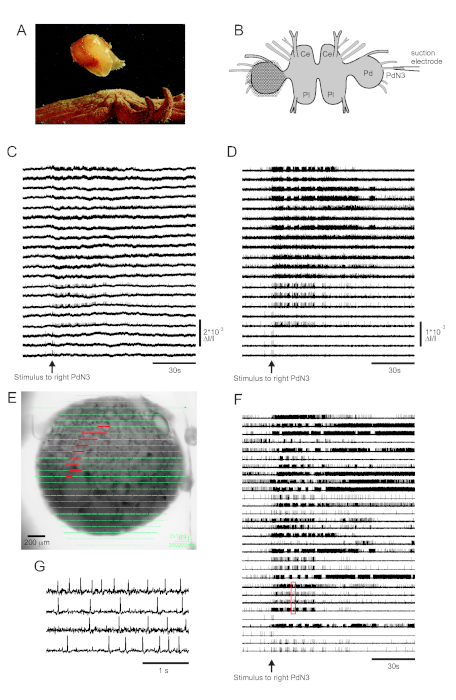
Figure 3: Results from Tritonia diomedea, illustrating raw, filtered, and spike-sorted data. (A) Tritonia escaping from the predatory sea star Pycnopodia helianthoides through its swim, which consists of alternating dorsal and ventral flexions of the body. (B) Schematic of the imaging setup. Ce=cerebral lobe of the cerebropleural ganglion; Pl = pleural lobe of the cerebropleural ganglion; Pd = pedal ganglion. (C) Raw data from 20 photodiodes, displaying activity in the left pedal ganglion to stimulation of the contralateral PdN3 (stimulus indicated by the arrow). (D) Filtered data from the same diodes as in C (5 and 100 Hz bandpass Butterworth filter). (E) Imaging software output in which compressed traces collected by all 464 diodes are superimposed topographically over an image of the preparation. The positions of the 20 diodes whose traces are shown in C and D are highlighted in red. (F) Thirty selected single-neuron traces generated by spike-sorting via ICA. (G) An expanded view of four single-neuron traces, corresponding to the red box in F, displays their action potentials at higher temporal resolution. Please click here to view a larger version of this figure.
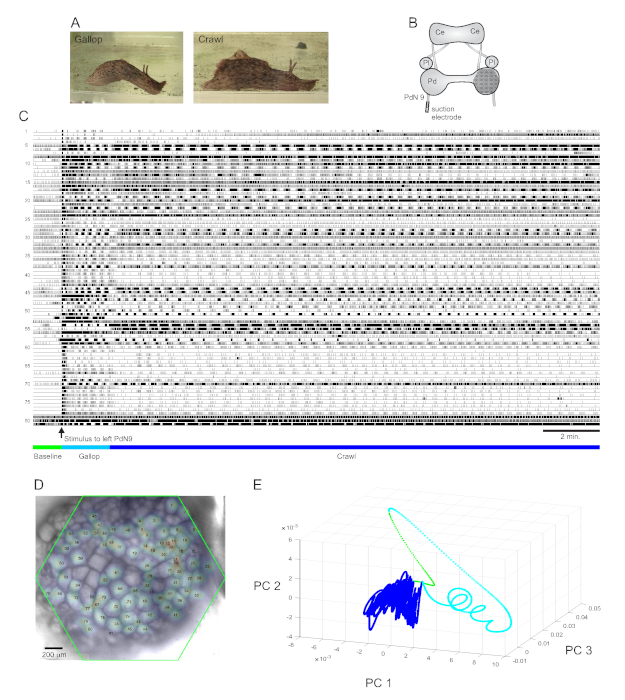
Figure 4: Results from Aplysia californica, illustrating long-duration recording, signal mapping, and dimensionality reduction. (A) The two phases of Aplysia’s sequential escape motor program, the gallop and the crawl. (B) Schematic of the imaging setup. Ce = cerebral ganglion; Pl = pleural ganglion; Pd = pedal ganglion. (C) A 20 min recording of 81 neurons in the right pedal ganglion responding to a stimulus to the contralateral PdN9 (indicated by the arrow). Green, cyan, and dark blue bars below the traces indicate the pre-stimulus period, the gallop, and the crawl phases of the escape motor program, respectively. (D) An image of the preparation with mapped probabilistic Gaussian distributions of the locations of all 81 neuronal signal sources identified by ICA. The green outline represents the position of the hexagonal face of the PDA with respect to the ganglion. The numbers on each Gaussian correspond to the trace numbers in C. (E) Dimensionality reduction using principal component analysis plotting the first three principal components against each other over the course of the 20 min file. The pre-stimulatory baseline, gallop, and crawl epochs are shown in green, cyan, and dark blue, respectively. See Videos 1-3 for animations of neuronal firing corresponding to this recording. Please click here to view a larger version of this figure.
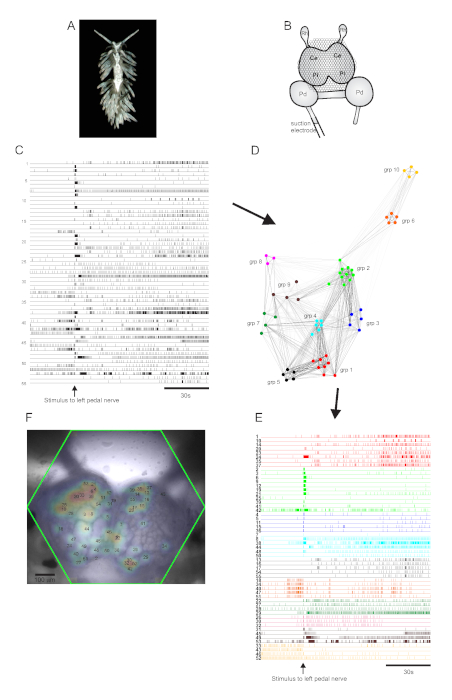
Figure 5: Results from Berghia stephanieae, a new species for neuroscience, illustrating network graphing, functional clustering, and bilateral signal mapping. (A) A Berghia specimen. (B) Schematic of the imaging setup. Ce = cerebral lobe of the cerebropleural ganglion; Pl = pleural lobe of the cerebropleural ganglion; Pd = pedal ganglion; Rh = rhinophore ganglion. (C) Traces displaying the spontaneous and stimulus-evoked activity of 55 bilateral neurons in the cerebropleural ganglia (delivery of the stimulus is indicated by the arrow). (D) A network graph displaying the ten functional ensembles, each assigned a unique color, identified through consensus clustering. Nodes in this plot represent neurons, where distance in network space represents the degree of firing correlation within and between ensembles. (E) The traces in C are rearranged and color-coded (following the color scheme of D) into functional ensembles. (F) An image of the preparation showing the mapped locations of the signals from every recorded neuron, and the trace numbers in C and E to which they correspond. Please click here to view a larger version of this figure.
Video 1: Animation of the full, 20-minute Aplysia escape locomotor program. The opacity of the white shapes overlying 81 individual neurons in the right pedal ganglion (left panel) was driven by the corresponding neuronal traces (right panel) and varied linearly as a function of average spike rate (binned per every 0.61 s of real time in the recording). For each neuron, full opacity was normalized to its maximum firing rate over the duration of the recording. One second of elapsed time in the video represents 12.2 s of real time. The scale bar corresponds to real time, with the green, cyan, and dark blue lines below the traces indicating the pre-stimulus baseline, gallop, and crawl phases of the escape locomotor program, respectively. The yellow boxes around the gallop phase and a portion of the crawl phase indicate the recording excerpts used to generate the animations in Videos 2 and 3. Please click here to view this video. (Right-click to download.)
Video 2: Animation of the gallop phase of the Aplysia escape locomotor program. Consensus clustering was performed on all 81 recorded neurons in just the gallop phase of the motor program to derive the functional ensembles, using the approach and software described and made available in ref.12. Neuronal ensembles exhibiting largely tonic or irregular firing patterns during this phase of the escape program were omitted from this video. The action potentials of the neurons belonging to the black and olive-green ensembles can be heard in the audio track of the video, with the corresponding neurons and traces highlighted. Average spike rates were normalized as in Video 1 and with equivalent time binning; 1 s of elapsed time in the video corresponds to 6.1 s of real time. Please click here to view this video. (Right-click to download.)
Video 3: Animation of the crawl phase of the Aplysia escape locomotor program. Consensus clustering was performed on all 81 recorded neurons in just the crawl phase of the motor program to derive the functional ensembles. Ensembles exhibiting largely tonic or irregular firing patterns during this phase of the motor program were omitted from this video. Average spike rates were normalized as in Videos 1 and 2 and with equivalent time binning; 1 s of elapsed time in the video corresponds to approximately 12.2 s of real time. Please click here to view this video. (Right-click to download.)
