After electrophysiological recordings, immunofluorescence is used to define the area infected by the AAV injection and to validate the expression of cofilinS3D (Figure 2). Immunostaining can be performed using a methodology similar to what has been described previously29,37,38,39. The AAV expresses an inactive form of cofilin fused with a hemagglutinin (HA)-Tag (cofilinS3D-HA), which is detected by immunofluorescence using an anti-HA antibody and a secondary antibody (Alexa Fluor 488). The infected excitatory neurons (here, targeted with a calcium/calmodulin-dependent protein kinase II alpha [CamKIIα] promoter controlling the expression of the transgene contained in the AAV) are stained with the anti-HA antibody. A successful infection is indicated by the staining of the neurons in the motor cortex surrounding the injection site (Figure 2A,B). In this representative example, the cerebral cortex of the other hemisphere did not show any noticeable staining. Nonetheless, given that excitatory neurons can project to distant brain areas, staining in the contralateral hemisphere is not necessarily an indication of unsuccessful injection. Higher magnification of the infected area showed staining of cell bodies and projections, confirming that only specific cells of the targeted cortical area were infected (Figure 2C).
Co-staining with markers of excitatory neurons (e.g., vesicular glutamate transporter 1, CaMKIIα) can also be performed to validate cell type-specificity. Alternatively, co-staining with markers of inhibitory neurons or astrocytes can be performed in case these cells are targeted using different promoters. Co-staining of cofilinS3D-HA and CaMKIIα was also performed in the same animal for an area more posterior to the injection site that still showed anti-HA staining in the motor cortex (Figure 2D). The higher magnification image of the area shows cells clearly expressing cofilinS3D-HA (Alexa Fluor 488, Figure 2E) and CaMKIIα (Alexa Fluor 568, Figure 2F). The superposition of the cofilinS3D-HA and CaMKIIα staining reveals that most (if not all) cells stained for cofilinS3D-HA are also positive for CaMKIIα (Figure 2G). This observation supports the specificity of the infection for excitatory neurons.
To assess the impact of cofilin manipulation on ECoG activity, ECoG and EMG signals are used to perform a visual identification of vigilance states (wakefulness, slow wave sleep, paradoxical sleep). This is done on 4-s epochs because of the rapid change in vigilance state in the mouse2, and here, for a full 24-h recording. Standard analyses include computation of sleep architecture and spectral analysis variables, as conducted previously for different datasets11,12,13,28,34. In particular, spectral analysis of the ECoG signal of the different states will index state composition and quality. To remove differences that could arise, for instance, from different depths of the electrodes, spectral analysis data can be expressed relative to the total power of all states of a given animal (Figure 3A). Given the very low relative amplitude of ECoG activity in higher frequencies, relative power spectra for wakefulness, slow wave sleep, and paradoxical sleep have been log-transformed to more adequately visualize and simultaneously compare the activity in low and high frequencies. This analysis indicates state-specific differences in spectral activity under conditions of cofilin inactivation (Figure 3B). More precisely, these preliminary findings combining male and female mice point out that cofilin inactivation significantly increases spectral power in fast frequencies (14-30 Hz) during wakefulness and in slow frequencies (1-4 Hz) during paradoxical sleep, while leaving ECoG activity during slow-wave sleep mainly unaffected. In addition, cofilin inactivation appears to increase inter-mouse variability in ECoG activity (particularly noticeable from error bars for wakefulness in Figure 3B).
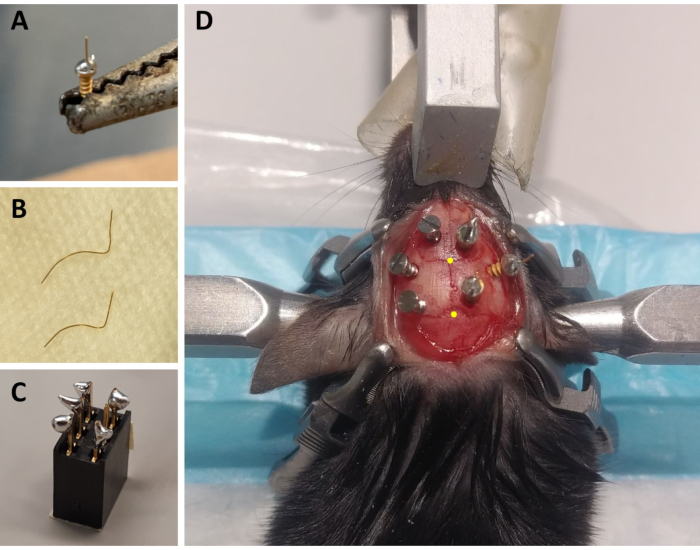
Figure 1: Preparation of ECoG/EMG montage components and representative example of ECoG electrode placement. (A) An ECoG electrode: a 4 mm long, 0.2 mm diameter gold wire (non-insulated) is fused on the head of a gold-covered screw (1.9 mm head diameter, 1.14 mm thread major diameter, 3.6 mm total length) using lead-free solder. (B) EMG electrodes: two gold wires (1.5 and 2 cm) are curved to embrace the curve of the skull up to the neck muscle, and the other end is kept straight to be soldered to the connector. (C) A 6-channel connector: lead-free solder is added to 5 of the 6 metal pins (omitting one in the middle) of the connector (5 mm x 8 mm x 8 mm + 3 mm metal pins). The top of the connector is covered with tape to avoid litter/water infiltration. (D) Example of the positioning of the three maintenance screws on the skull of the left hemisphere and of the three ECoG electrodes (including a reference electrode) on the right hemisphere. The precise stereotaxic coordinates of the ECoG electrodes are indicated in steps 2.6 and 3.2 and have been calculated according to the location of the bregma and lambda (which are indicated by the yellow dots). Abbreviations: ECoG = electrocorticographic; EMG = electromyographic. Please click here to view a larger version of this figure.
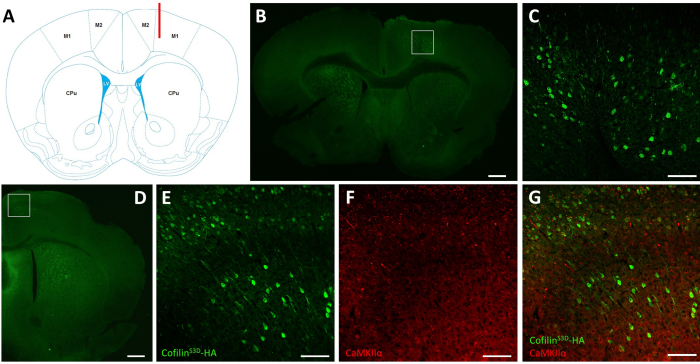
Figure 2: Representative immunostaining to define the AAV-infected area and cell type. (A) Schematic representation showing the injection site of the coronal slice presented in panel B. The position is 1.1 mm anterior to the bregma, and the cannula (shown in red) was targeted to layers V of the right primary motor cortex (M1). Representation modified from Franklin and Paxinos40. (B) Immunostaining of HA to detect cofilinS3D-HA expression in neurons shown for a coronal slice of the full brain located approximately 1.1 mm anterior to the bregma. The infected area mainly localizes to layers V and VI (infragranular layers) of the right primary and secondary motor cortices (M1 and M2). Scale bar = 500 µm. The square represents the area shown in C. (C) Higher magnification of the infected area showing staining of infected cells and confirming expression of cofilinS3D-HA in deeper layers of the motor cortex. Scale bar = 100 µm. (D) Co-immunostaining of HA and CaMKIIα to assess cell type-specificity shown for a coronal slice of the right hemisphere located approximately 0.5 mm anterior to bregma and therefore, posterior to the injection site (same mouse as in panels B and C). The infected area localizes to motor cortices (M1 and mainly M2). Scale bar = 500 µm. The square represents the area shown in E, F, and G. (E) Higher magnification of the infected area showing staining of infected cells and confirming expression of cofilinS3D-HA. Scale bar = 100 µm. (F) Higher magnification of the infected area showing staining of CaMKIIα-positive cells. Scale bar = 100 µm. (G) Higher magnification of the infected area showing co-labeling of cofilinS3D-HA and CaMKIIα, confirming that infected cells are CaMKIIα-positive. Scale bar = 100 µm. Abbreviations: AAV = adeno-associated virus; M1 = primary motor cortex; M2 = secondary motor cortex; CPu = caudate putamen (striatum); LV = lateral ventricle; HA= hemagglutinin; CamKIIα = calcium/calmodulin-dependent protein kinase II alpha. Please click here to view a larger version of this figure.
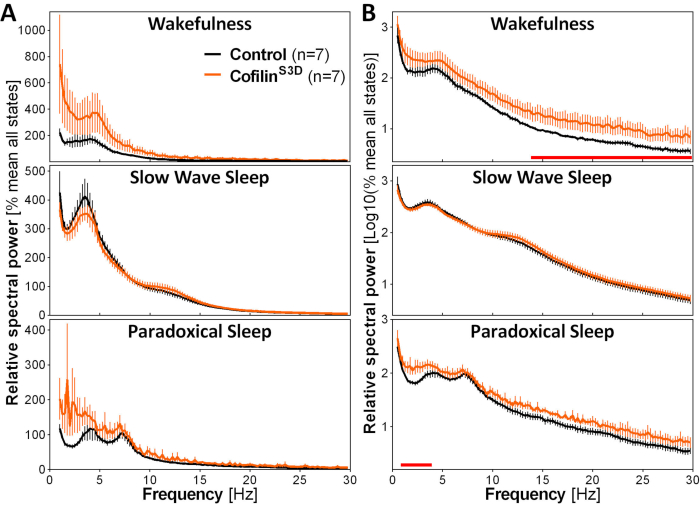
Figure 3: Representative power spectra for wakefulness, slow wave sleep, and paradoxical sleep obtained after viral manipulation of cofilin function. Male (n = 5 per group) and female (n = 2 per group) mice injected with AAV9-CaMKIIα0.4-cofilinS3D-HA (viral titer 2.58 × 1013 GC/mL) or with a control AAV (AAV9-CaMKIIα0.4-eGFP 1.25 × 1013 GC/mL; half of the test titer to control for the enhanced signal of this control AAV) in layer V of the motor cortex were recorded for 24 h, and the electrocorticographic signal was subjected to spectral analysis (fast Fourier Transform to calculate spectral power between 0.5 and 30 Hz with a 0.25-Hz resolution). (A) Power spectra during the three vigilance states expressed relative to total power of all states. (B) Relative power spectra log-transformed to more adequately represent group differences in higher frequencies. The suppression of cofilin activity in the motor cortex using AAV9-CaMKIIα0.4-cofilinS3D-HA significantly increases electrocorticographic activity in the beta range (14-30 Hz) during wakefulness, and in the delta range (1-4 Hz) during paradoxical sleep in comparison to control injections (red lines above x axes indicate Mann-Whitney U-test on frequency band power p < 0.05). Abbreviations: AAV = adeno-associated virus GC = genome copies; HA= hemagglutinin; CamKIIα = calcium/calmodulin-dependent protein kinase II alpha; eGFP = enhanced green fluorescent protein. Please click here to view a larger version of this figure.
