We describe here how the cone-enriched culture system can be used to identify novel cone protecting proteins. We used this protocol to screen a normalized cDNA library made of choroid and retinal pigmented epithelium from 400 eyes of 8 weeks old Long-Evans rats43.
This library contains 6.0 x 106 independent colonies forming units (CFUs) and has an average cloned insert size of 2.1 kilobase (kb), with greater than 99% of recombinant clones. Pools of 100 clones from that library were transiently transfected (0.1 µg of plasmid DNA) into COS-1 cells and the conditioned medium (CM) of COS-1 were harvested after incubation for 48 hours in DMEM without serum. Membranes or exosomes were not removed by ultracentrifugation. Fifty microliters of each CM were added to 4 wells of two 96-well plates: one seeded at 2 x 105 cells/cm2, the other one at 4 x 105 cells/cm2. CM from COS-1 cells transfected with the empty vector pcDNA3.1 used to construct the library was used as negative control (Table 1). A total of 2,112 sets of 100 clones corresponding to 211,200 individual clones were evaluated in four culture wells and for two seeding conditions of cone-enriched cultures. The two conditions correspond to two slightly different inoculation densities which makes it possible to assess the protective activity more precisely, for a total of 1,689,600 culture wells.
Among the 42 pools of clones with a ratio greater than 2, pools 0080 and 0073 have viability ratio 16 and 14 times higher after 7 days of culture than the negative control, pcDNA3.1 (Supplementary Figure 1). This analysis is essential to identify the pools of interest. Each selected pool of 100 clones was subdivided into 16 sets of 10 clones from their glycerol stock. These sub-pools were prepared and tested according to the same method in a second round of screening (i.e., a total of 3,200 culture wells). The sub-pool 0073-09 gave the strongest viability ratio (Supplementary Figure 2A) and was subdivided to produce 16 individual clones that were tested in a third round of screening on cone-enriched cultures. The clone 0073-09-37 clone clearly stands out from the others with a viability ratio equal to 2.5 (Supplementary Figure 2B). The y axis has a different scale even if the seeding density was the same than in Supplementary figure 2A. We have seen this commonly when the assays are repeated weekly for months. After analysis, these results confirm that clone 0073-09-37 has a robust and reproducible effect on cone survival. The test was repeated independently (Figure 3A), and the insert of 1.8 kb was sequenced (Figure 3B).
A bioinformatic analysis revealed that the clone 0073-09-37, that we named epithelium-derived cone viability factor (EdCVF), contains three open reading frames (ORFs), the one the most upstream (ORF1) encodes for 84 residues of the C-terminal part of the rat protein zinc finger protein-180 (ZFP180, NP_653358) of 727 amino acids44. The other two ORFs (ORF2 and ORF3) are much less well conserved in mice and absent in other mammals. When tested independently, only ORF1 exerts a protective effect on the cones (Supplementary Figure 3). ORF1 was produced as a glutathione S-transferase (GST) fusion protein (Figure 4A). The EdCVF protein was purified and the GST tag removed (Figure 4B). EdCVF is able to prevent cone degeneration in the cone-enriched culture system (Figure 4C).
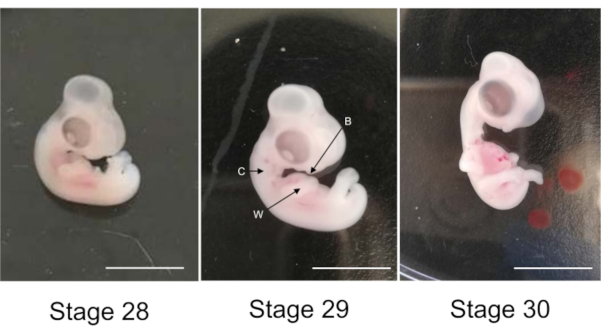
Figure 1: Chicken embryos at stages 28th, 29th and 30th of development. Arrows W: wing, C: collar and B: bill. Scale bar 1 cm. Please click here to view a larger version of this figure.
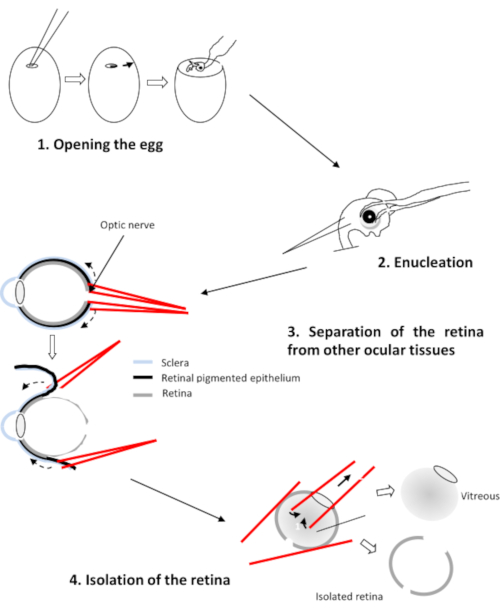
Figure 2: Dissection of the retina of the chicken embryo Please click here to view a larger version of this figure.
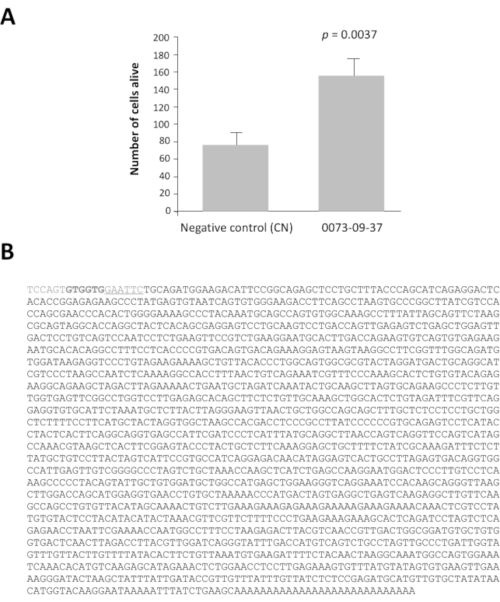
Figure 3: Epithelium-derived cone viability factor (EdCVF), clone 0073-09-37. A. Increased viability on cone-enriched culture. B. The sequence of the cDNA clone 0073-09-37. The underlined sequence GAATTC is the EcoRI restriction site used to construct the library. The two codons GTG in bold are uncommon translation initiation sites for EdCVF originating from the vector. Statistical analysis by Student's test. Please click here to view a larger version of this figure.
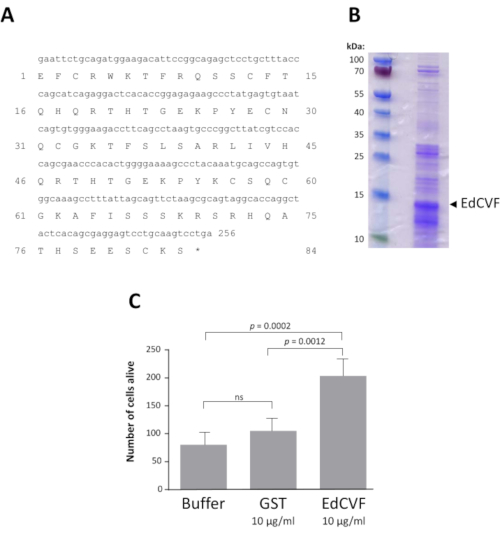
Figure 4: Recombinant EdCVF activity. A. Sequence of EdCVF in the fusion with glutathione S-transferase (GST). B. The purified recombinant EdCVF protein. C. Trophic activity of GST-EdCVF on cone in culture. Statistical analysis using Tukey's test. Please click here to view a larger version of this figure.
Supplementary Figure 1: Ratio of the average cell number for a cDNA pool and the negative control, the conditioned medium of COS-1 cells transfected with the empty vector pcDNA3.1 during the first round of screening. Please click here to download this file.
Supplementary Figure 2: Number of cells alive. A. The second round of screening with sub-pools of 0073 pool. B. The third round of screening with isolated clones. CN: the conditioned medium of COS-1 cells transfected with the empty vector, pcDNA3.1. Please click here to download this file.
Supplementary Figure 3: Trophic activity of the three open reading frames of the isolated clone 0073-09-37. Statistical analysis using Dunnett's test. Please click here to download this file.
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | |
| A | 1 | 1 | 1 | 1 | 2 | NC | 2 | 2 | 2 | 3 | 3 | NC |
| B | NC | 3 | 3 | 4 | 4 | 4 | NC | 4 | 5 | 5 | 5 | 5 |
| C | 6 | NC | 6 | 6 | 6 | 7 | 7 | NC | 7 | 7 | 8 | 8 |
| D | 8 | 8 | NC | 9 | 9 | 9 | 9 | 10 | NC | 10 | 10 | 10 |
| E | 11 | 11 | 11 | NC | 11 | 12 | 12 | 12 | 12 | NC | 13 | 13 |
| F | 13 | 13 | 14 | 14 | NC | 14 | 14 | 15 | 15 | 15 | NC | 15 |
| G | 16 | 16 | 16 | 16 | 17 | NC | 17 | 17 | 17 | 18 | 18 | NC |
| H | 18 | 18 | 19 | 19 | 19 | 19 | NC | 20 | 20 | 20 | NC | 20 |
| NC negative controls | ||||||||||||
| 1-20 positions of the pools tested in quadruplicate | ||||||||||||
Table 1: Plan of the 96-well plate for high content screening
