The conditioned medium was collected from lentivirus of EcoHIV-EGFP infected 293FT cells. Next, it was concentrated and titered, then stereotaxically injected into the brain (cortical region) of F344/N rats. Seven days post-injection, rats were sacrificed and images were taken from coronal brain slices ranging from bregma 5.64 mm to bregma -4.68 mm. In Figure 1A, there are significant EcoHIV-EGFP signals throughout the brain, especially in the cortex and the hippocampal dentate gyrus. Furthermore, dual-labeling with Iba1and EcoHIV-EGFP probes provided strong evidence that microglia were the predominant cell type harboring EcoHIV expression in the brain (Figure 1B). Notably, the distribution pattern of EcoHIV-EGFP is consistent with the relative regional concentrations of microglia in the rat brain (i.e., cortex and dentate gyrus of the hippocampus).
In a subsequent study, we validated the utility of EcoHIV infection in rats to model key aspects of HAND. Using the protocol detailed above, F344/N rats were stereotaxically injected with either EcoHIV-EGFP or saline. First, eight weeks post-infection, temporal processing, a potential elemental dimension of HAND12, was evaluated using visual gap prepulse inhibition (Figure 2). EcoHIV animals exhibited a relative insensitivity to the manipulation of interstimulus interval (ISI), evidenced by a relatively flatter ISI function compared to saline controls. Specifically, significant differences in the slope of the ISI function from the 50 ms ISI to the 200 ms ISI were observed (Semilog Line-X is Log, Y is Linear, R2s ≥ 0.99; F(1,2)=642.9, p≤0.001). Second, ballistic labeling was used to investigate the impact of EcoHIV-EGFP injections on the morphology of dendritic spines in medium spiny neurons (MSN) of the nucleus accumbens (NAc; Figure 3); parameters which can be utilized to draw inferences about synaptic function13. EcoHIV rats displayed profound alterations in dendritic spine morphology, evidenced by an increased relative frequency of shorter dendritic spines (Genotype x Bin Interaction, F(16, 218) = 4.3, p ≤ 0.001) with an increased head diameter (Genotype x Bin Interaction, F(12, 96) = 18.7, p ≤ 0.001) and increased neck diameter (Genotype x Bin Interaction, F(15, 120) = 16.3, p ≤ 0.001) relative to control animals. Detailed methodology for the assessment of temporal processing14 and ballistic labeling13 have been previously reported.
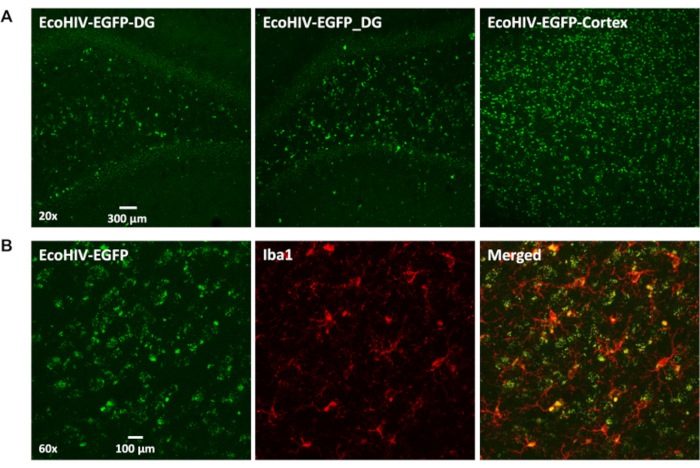
Figure 1. The EcoHIV-EGFP infected cells distributed in rat brain. (A) The representative confocal images (20x) of EcoHIV-EGFP expression in hippocampal dentate gyrus or cortex regions at 7 days after injection. (B) The representative confocal images (60X) of co-localization of Iba1 immunostaining with EcoHIV-EGFP infected cells at 7 days after injection. Please click here to view a larger version of this figure.
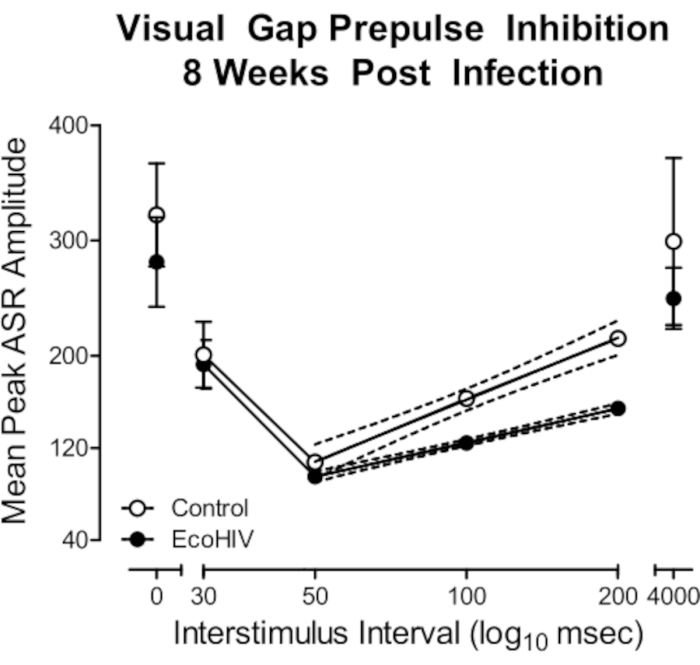
Figure 2. EcoHIV infection induced prominent neurocognitive deficits in temporal processing. Visual gap prepulse inhibition was conducted eight weeks after stereotaxic injections of either EcoHIV or saline. EcoHIV infection induced prominent alterations in temporal processing evidenced by the relative insensitivity to the manipulation of interstimulus interval relative to control rats. Detailed methodology described in McLaurin et al.13. Please click here to view a larger version of this figure.
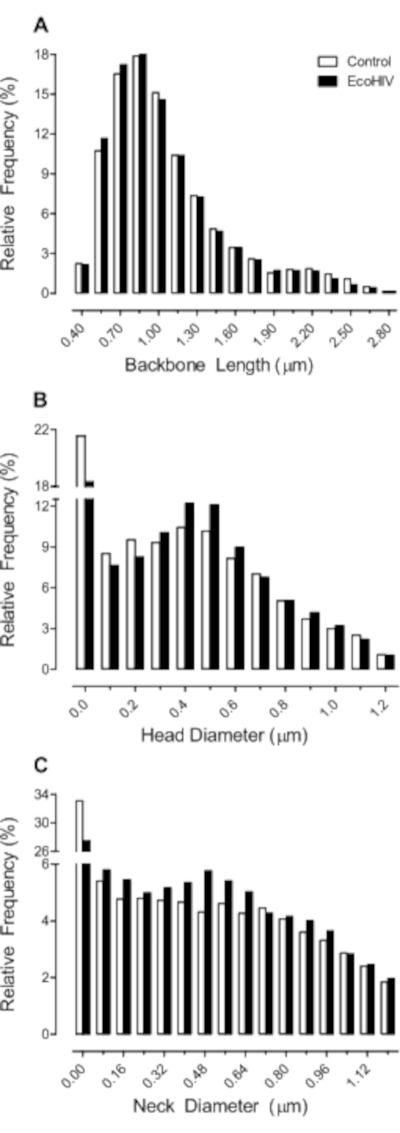
Figure 3. Infectivity with EcoHIV-EGFP altered the morphological parameters of dendritic spines, supporting profound synaptic dysfunction. EcoHIV rats displayed profound alterations in dendritic spine morphology, evidenced by an increased relative frequency of shorter dendritic spines (A) with an increased head diameter (B) and increased neck diameter (C) relative to control animals. Please click here to view a larger version of this figure.
