We have extensively illustrated that this protocol is a valid assay for determining the rate of import into functional and intact isolated skeletal muscle mitochondria. In comparison to untreated conditions, the import of typical precursor proteins such as malate dehydrogenase (MDH) into the matrix is sensitive to membrane potential because it can be inhibited by valinomycin, a respiratory chain uncoupler (Figure 2A). Import is also impeded when mitochondrial inner and outer membranes are solubilized in the presence of the detergent Triton X-100. The import process is sensitive to the presence of external ATP, which serves to unfold precursor proteins for translocation across the membranes, and is tightly controlled by the rate of respiration and ATP provision (data not shown20). Distinct differences in the import are also observed between intermyofibrillar and subsarcolemmal mitochondrial fractions, in part due to variations in protein import machinery expression, as well as the respiratory rate between these mitochondria (Figure 2B).
Mitochondria in skeletal muscle are highly dynamic organelles that respond readily to changes in energy demand. Mitochondrial content in the muscle increases following periods of chronic exercise or in response to electrical stimulation-induced contractile activity (see for review21). For example, 7 days of chronic contractile activity of rat skeletal muscle enhances import into the OM and matrix by 1.6-fold and 1.4-fold, respectively22 (Figure 3A, 3B). These changes in mitochondrial content are brought about, in part, by alterations in the capacity of the mitochondrial protein import system. Indeed, a close relationship between the import rate of precursor proteins and a good estimate of mitochondrial content, as measured by the complex IV marker cytochrome oxidase under control or denervated conditions, can be illustrated23 (Figure 3C).
The adaptability of the import system in muscle to alterations in contractile activity suggests that exercise could be used as a treatment to resolve defects in the import pathway if identified. During the investigation of mitochondrially-mediated apoptosis using Bax-Bak double knockout animals, we noticed that the reduced mitochondrial content in the muscles of these experimental animals was accompanied by a decrease in the import of precursor proteins into the matrix. We then investigated the possibility that exercise could restore this import capacity. Indeed, following 6 weeks of voluntary wheel run training, protein import was restored to control levels in the knockout animals24, illustrating the adaptive plasticity of the import pathway to rescue mitochondrial content and function (Figure 4).
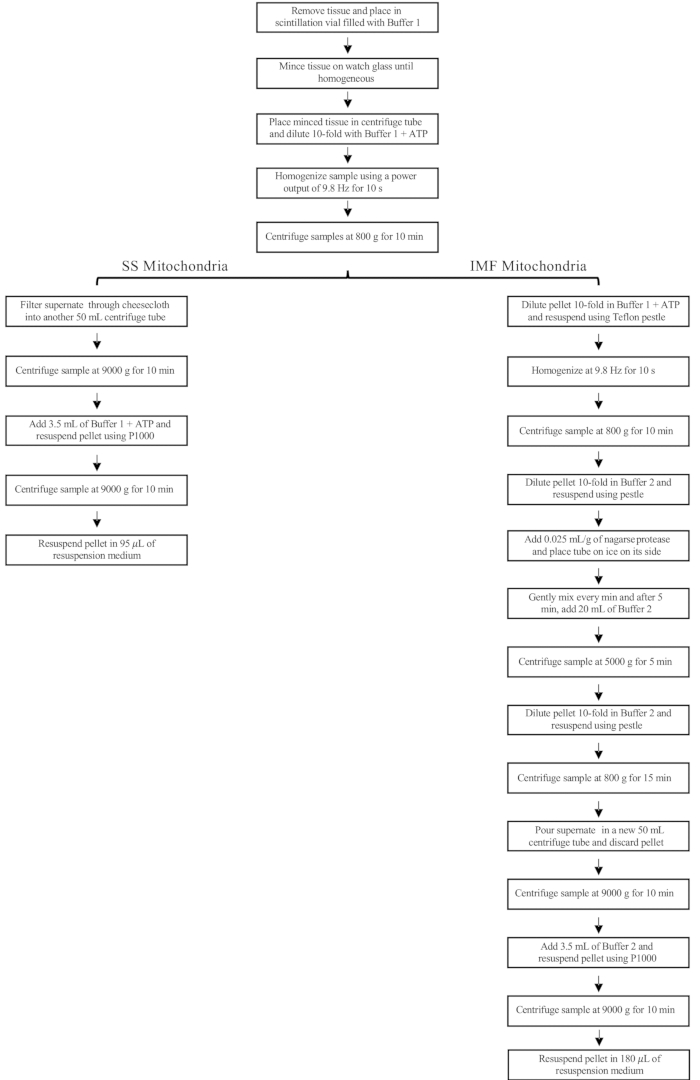
Figure 1: Schematic of the workflow for isolating SS and IMF mitochondria from skeletal muscle based on the previous work8. This protocol allows the isolation of functional mitochondria based on their geographic location within skeletal muscle. SS mitochondria are more rapidly and easily liberated, while the IMF mitochondria require a further digestion step with protease to untangle them from the myofibrils. Note that the isolation of these subfractions can be done in tandem. An updated, similar procedure has recently been published25. Please click here to view a larger version of this figure.
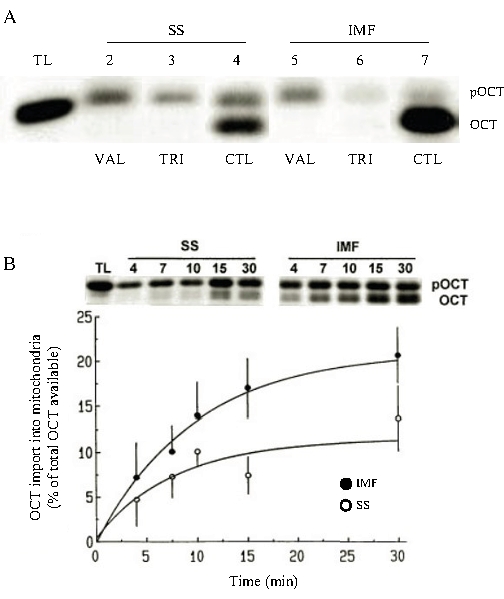
Figure 2: Protein import into the mitochondrial matrix. (A) Normal rates of import for MDH are shown in SS and IMF mitochondria (lanes 4 and 7) and the translation product of precursor MDH (lane 1). The lower band represents the imported mature MDH. Addition of valinomycin inhibits MDH protein import into the matrix of SS and IMF mitochondria, as this uncoupler dissipates the membrane potential (lanes 2 and 5). Triton-X is a detergent that solubilizes the inner membrane, thereby inhibiting MDH import into these subfractions (lanes 3 and 6). Protein import was carried out for increasing time durations, 4 min, 7 min, 10 min, 15 min, and 30 min. These data illustrate that import is a time-dependent process and also that SS and IMF mitochondria have different rates or capacities for import (B). TL, translation lane; VAL, valinomycin; TRI, Triton-X; CTL, control; SS, subsarcolemmal; IMF, intermyofibrillar. This figure was modified from Takahashi M & Hood DA20. Please click here to view a larger version of this figure.
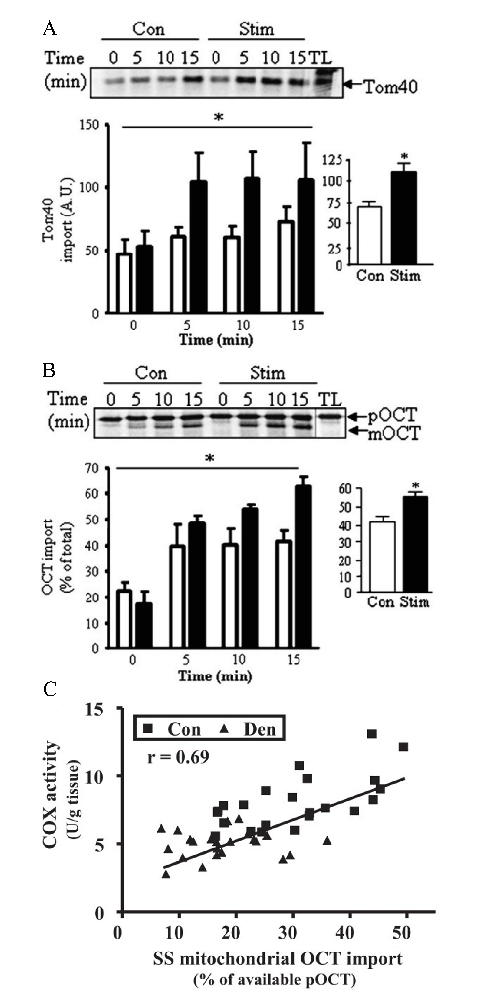
Figure 3: Protein import is adaptable and tightly linked to estimates of mitochondrial content. Sprague Dawley rats were subjected to electrical stimulation to induce contractile activity, a model of exercise training. (A) Tom40 import into the OM was 1.6-fold higher in muscle from chronically stimulated animals compared to controls at any given time point. (B) OCT import into the mitochondrial matrix was increased at every time point of incubation, and overall, this resulted in a 1.4-fold increase in mitochondria from chronically stimulated muscle. (C) Import is positively correlated with an index of mitochondrial content, as assessed by COX activity, r = 0.69. These measurements were taken from animals that were subjected to denervation, which has been shown to decrease mitochondrial content and the rate of import. Con, control; Den, denervated; Stim, stimulated; TL, translation lane; * p < 0.05. Figures 3A and 3B were adapted from Joseph A-M & Hood DA22 and 3C from Singh B & Hood DA23. Please click here to view a larger version of this figure.
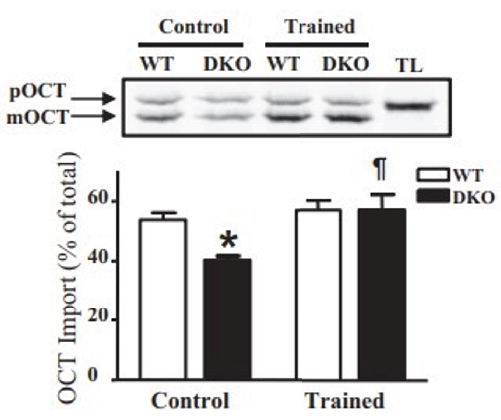
Figure 4: Training rescues import defects in vivo. Bax/Bak double knockout animals exhibit reduced protein import into the mitochondrial matrix by 37%. Six weeks of voluntary wheel running rescued the import defect to control levels. WT, wildtype; DKO, double knockout; TL, translation lane; * p < 0.05 main effect of genotype; ¶ p < 0.05 main effect of training. This figure was adapted from Zhang Y et al.24. Please click here to view a larger version of this figure.
| Buffer 1: | Buffer 1 + ATP: | Buffer 2: | Resuspension medium: | Nagarse protease |
| 100 mM KCl | 100 mM KCl | 100 mM KCl | 100 mM KCl | 10 mg/mL in Buffer 2 |
| 5 mM MgSO4 | 5 mM MgSO4 | 5 mM MgSO4 | 10 mM MOPS | NOTE: Make fresh and keep on ice |
| 5 mM EDTA | 5 mM EDTA | 5 mM EGTA | 0.2% BSA | |
| 50 mM Tris | 50 mM Tris | 50 mM Tris | ||
| 1 mM ATP | 1mM ATP |
Table 1: Buffers and resuspension media.
| % of the reaction volume | 1 Reaction mix | |
| Reticulocyte Lysate | 64.10% | 11.8 µL |
| Amino acids (-methionine) | 2.20% | 0.4 µL |
| Sterile H2O | 21.60% | 3.97 µL |
| 35S-methionine | 7.20% | 1.33 µL |
| mRNA | 5.40% | 1.0 µL |
| Total Volume | 18.5 µL | |
| NOTE: | ||
| 1) Add 35S-methionine last; | ||
| 2) Thaw the lysate slowly on ice, and limit freeze/thaw cycles to two. It is recommended that the lysate be aliquoted upon arrival | ||
| 3) The volume of mRNA can be adjusted to optimize translational efficiency by altering the sterile H2O volume accordingly. | ||
Table 2: Translation reaction mix
