After isolation and dissociation of the AL, the obtained single cells are seeded in ECM and grown in PitOM (Figure 1, Table 1). Figure 3A displays the cell culture and density at seeding (Day 0). Some small debris may be present (Figure 3A, white arrowheads), but will disappear at passaging. Fourteen days after seeding, the AL-derived organoids are fully developed (Figure 3A). The organoids exhibit a cystic morphology, with an epithelial layer that encloses a lumen. At this stage, the organoids reach a diameter of 500 µm and have to be passaged. Figure 3B shows the AL-derived organoid culture after passaging at the indicated time following re-seeding of the dissociated organoid fragments.
Occasionally, one or more dense structures may appear in the organoid culture (Figure 3A, Unfavorable). When passaging, dense organoids tend to take over, ending up in cultures with only dense structures after a couple of passages (Figure 3B, Unfavorable). Therefore, it is recommended not to proceed with wells that contain dense organoids (passage 0). Alternatively, dense organoids can be discarded by sedimentation, which leaves the cystic organoids to continue with. The origin of these dense organoids is at present unclear, but they show a less pronounced pituitary nature18. If organoids do not, or less efficiently regrow after passaging, dissociation procedures need to be optimized. In particular, one must pay attention not to dissociate too harsh; the organoids must be split up to fragments, not to single cells (Figure 3B, Day 0, inset).
Immunofluorescence staining analysis confirms the epithelial character of the AL-derived organoids, as they express the epithelial markers E-cadherin (E-Cad) and cytokeratin 8/18 (CK8/18; Figure 3C), which, moreover, have been described as stem cell markers in the pituitary18. The stemness nature of the organoids is additionally demonstrated by SOX2 and TROP2 expression, both of which were also identified as pituitary stem cell markers (Figure 3C)14,18. LHX3, a transcription factor specifically expressed in the (early-developing) pituitary, validates the organoids' pituitary phenotype (Figure 3C). Some of the organoid-constituting cells are in a proliferative state, expressing the proliferation marker Ki67 (Figure 3C).
Further exploration and validation of the pituitary (stemness) phenotype of the AL-derived organoids is performed with reverse transcription-quantitative PCR (RT-qPCR). High expression of the stemness markers Sox2, Cdh1 (encoding E-Cad), Krt8, Krt18 and Trop2 is present in the organoids, clearly higher than in primary AL, indicating that the organoids enrich for the stem cells and thus represent the AL stem cell compartment, as previously described (Figure 3D)18. Notably, the developmental transcription factors Pitx1 and Pitx2 remain expressed after development in several hormonal cell types in the AL, and hence their high expression in the AL as well. The cultures robustly retain their stemness phenotype, as demonstrated by the sustained (high) expression of these markers after multiple passages (Figure 3D).
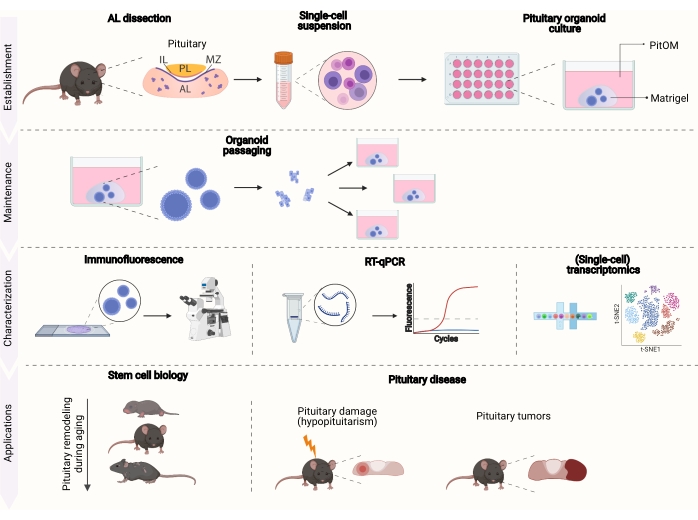
Figure 1: Overview of the establishment, maintenance, characterization, and application potential of organoids from healthy and diseased pituitary. AL, anterior lobe; IL, intermediate lobe; PL, posterior lobe; MZ, marginal zone; PitOM, pituitary organoid medium (created with BioRender.com). Stem cell niches in the AL are indicated in purple. Please click here to view a larger version of this figure.
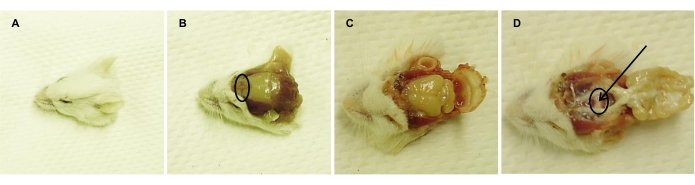
Figure 2: Isolation of the pituitary gland from adult euthanized mouse. Representative images consecutively taken following (A) decapitation, (B) removal of head skin (nose bridge is encircled), (C) opening of the cranium, and (D) removal of the brain, exposing the pituitary gland (encircled). Arrow points to the PL, which is discarded (together with the associated IL), leaving the AL for isolation and dissociation. Please click here to view a larger version of this figure.
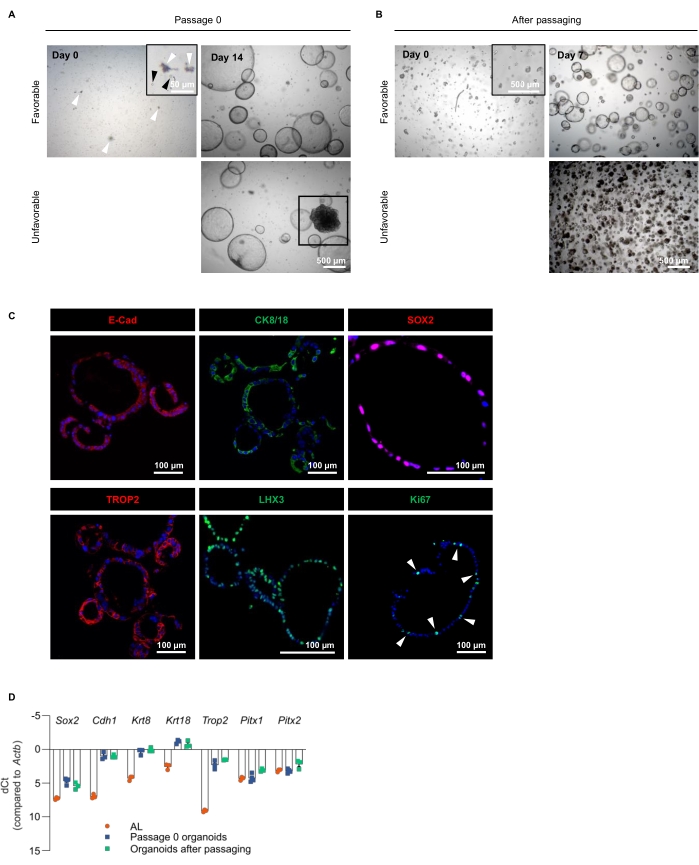
Figure 3: Establishment and validation of AL-derived organoids. (A) AL cell seeding and organoid development in PitOM at indicated days (passage 0). The top row shows favorable organoid growth, with only cystic structures developing. The bottom row shows unfavorable growth with a large dense structure appearing (boxed). White arrowheads indicate debris, black arrowheads indicate single cells (magnified in inset). (B) Organoid fragments (magnified in inset) seeded at passaging (Day 0) and regrowth of organoids as observed 7 days later. The top row shows favorable organoid regrowth, with only cystic structures growing. The bottom row shows unfavorable regrowth with dense organoids taking over the culture. (C) Immunofluorescence staining of E-Cad, SOX2, TROP2 (all red), CK8/18, LHX3 and Ki67 (all green) in AL-derived organoids. Nuclei are labeled with Hoechst33342 (blue). Arrowheads indicate Ki67+ cells. Scale bars are indicated. (D) Gene expression analysis of stemness markers (Sox2, Cdh1, Krt8, Krt18, Trop2), and developmental transcription factors (Pitx1, Pitx2) in primary AL and AL-derived organoids (Passage 0 means 14 days after cell seeding) determined by RT-qPCR (mean ± SEM). Data points represent biological replicates. Delta cycle threshold (dCT) values are shown, calculated using the formula: CT(gene of interest) – CT(housekeeping gene Actb). The more positive the dCT value (which is presented on the Y-axis below the zero X-axis), the lower the expression level of the gene of interest. The lower (or more negative) the dCT value, the higher the expression level14,18,21,22. Please click here to view a larger version of this figure.
| Pituitary organoid medium (PitOM) | |
| Component | Concentration |
| Advanced DMEM/F12 | |
| Hepes | 1% |
| Penicillin-Streptomycin | 1% |
| Glutamax | 1% |
| B-27 Supplement (50X), minus vitamin A | 1X |
| L-Glutamine (200 mM) | 2 mM |
| Recombinant Human FGF basic/FGF2/bFGF (157 aa) Protein | 20 ng/mL |
| Recombinant Human IGF-1 | 100 ng/mL |
| N-2 Supplement (100X) | 1X |
| N-acetyl-cysteine | 1.25 mM |
| Recombinant Human/Murine FGF-8b | 200 ng/mL |
| Recombinant Human FGF-10 | 100 ng/mL |
| A83-01 (activin receptor-like kinase 4/5/7 inhibitor) | 0.50 µM |
| Recombinant Mouse Sonic Hedgehog/Shh (C25II) N-Terminus | 100 ng/mL |
| Recombinant Human EGF Protein, CF | 50 ng/mL |
| SB202190 (p38 mitogen-activated protein kinase inhibitor) | 10 µM |
| Recombinant Human Noggin | 100 ng/mL |
| Cholera Toxin from Vibrio cholerae | 100 ng/mL |
| Recombinant Human R-Spondin-1 | 200 ng/mL |
| Recombinant Human IL-6 | 20 ng/mL |
Table 1. Composition of PitOM. PitOM is filtered through a 0.22 µm mesh filter and stored at 4 °C for a maximum of 2 weeks.
| Medium A | |
| Component | Quantity |
| DMEM, powder, high glucose | 13.38 g |
| HEPES | 5.96 g |
| Sodium-Pyruvate (C3H3NaO3) | 0.11 g |
| Penicillin G sodium salt | 35.00 mg |
| Streptomycin sulfate salt | 50.00 mg |
| Sodium Chloride (NaCl) | 0.50 g |
| Sodium Hydrogen Carbonate (NaHCO3) | 1.00 g |
| Albumin Bovine (cell culture grade) | 3.00 g |
| Sterile water | 1.00 L |
| Medium C | |
| Component | Quantity |
| Sodium Chloride (NaCl) | 7.50 g |
| Potassium Chloride (KCl) | 0.40 g |
| Sodium di-Hydrogen Phosphate 1-hydrate | 0.14 g |
| D-glucose | 1.00 g |
| HEPES | 4.76 g |
| Streptomycin sulfate salt | 50.00 mg |
| Penicillin G sodium salt | 35.00 mg |
| Phenol red | 10.00 mg |
| Albumin Bovine (cell culture grade) | 3.00 g |
| Sodium Hydrogen Carbonate (NaHCO3) | 1.00 g |
| Sterile water | 1.00 L |
| Medium B | |
| Component | Quantity |
| Titriplex III (Edetate disodium salt dihydrate) | 0.74 g |
| Medium C | 100 mL |
Table 2. Composition of medium A, B, and C. All media are filtered through a 0.22 µm mesh filter and stored at 4 °C for a maximum of 4 months. The pH of medium A and C must be adjusted to 7.3.
| Cryopreservation medium | |
| Component | Concentration |
| Advanced DMEM/F12 | 60% |
| FBS | 30% |
| DMSO | 10% |
Table 3. Composition of cryopreservation medium.
