Refer to Figure 1 for an illustration of the protocol. Ethical clearance was obtained from the University of Pretoria Health Sciences Ethics Committee (506/2018) for the withdrawal and use of human blood.
1. Gametocyte culture
NOTE: Prior to setting up the SMFA, a gametocyte culture was prepared at the University of Pretoria (see Reader et al.22 for the complete protocol).
- Prepare a gametocyte culture that consists of stage V gametocytes from the NF54 parasite strain.
- Ensure that the gametocytemia of the culture is between 1.5% and 2.5%, with a 50% hematocrit in A+ male serum, to which fresh red blood cells are added.
- Separate the culture into different flasks and add 2 µM of each compound for each respective treatment 48 h prior to conducting the SMFA. Leave the control group untreated.
- Assess the gametocyte culture shortly before conducting the SMFA to ensure the exflagellation of male gametes, with the presence of a 3:1 female:male ratio.
2. Artificial infection of mosquitoes through the SMFA
NOTE: Biosafety: infected mosquitoes should be housed in a biosafety level 2 (BSL2) facility with restricted access.
- Using a mouth aspirator, place 25 unfed female An. gambiae mosquitoes into a 350 mL feeding cup. Do the same for each treatment cup and label the cups clearly according to whether they are to be used as control or treatment groups. Choose the number of cups per treatment according to the number of technical replicates included.
NOTE: Colony mosquitoes between 5 and 7 days old are used in a typical transmission-blocking compound evaluation. Starving mosquitoes for 3-4 h or longer prior to blood feeding will facilitate the uptake of blood during SMFA. - Connect the glass feeder system to the water bath and maintain the temperature at 37 °C.
NOTE: The glass feeder consists of two arms, which are connected to the silicone tubing to which the water bath is connected (Figure 2). The hollow structure of the feeder allows water to circulate through and maintenance of the temperature of the blood. - Prepare cow intestine (or synthetic membrane) by rinsing it in tap water and cut it into pieces that are fitted for each feeder. Cover each feeder and fasten the membrane with an elastic band.
NOTE: No ethical clearance was needed for the intestine, as it was bought from a local butchery, where it is sold to the public for food preparation. - Place the infection cups underneath the feeders, with the membrane laying on top of the net of the cup.
- Add 1 mL of gametocyte-infected blood to the feeders of the control cups and gametocyte-infected blood with added compound to each corresponding compound feeder and cup.
- Leave the mosquitoes to feed for approximately 40 min with the feeders uncovered.
NOTE: Feedings take place under insectary conditions (25 °C, 80% relative humidity) in the dark. The diameter of a feeder is approximately 13 mm. - After feeding, remove the feeders from the cups, rinse the feeders, and treat the excess blood with hypochlorite.
- Remove the unfed mosquitoes from the cup by knocking all the mosquitoes down on ice (for 1-2 min) and separating the unfed mosquitoes from those that have taken a blood meal. Look for swollen and red abdomens (indicating blood) to distinguish the fed, fully engorged mosquitoes from the unfed ones (Figure 3).
- Place the infection cups in the biosafety chamber (Supplemental Figure S1) and provide each cup with a 10% sugar water pad, replacing the sugar water on alternate days for 8-10 days.
3. Preparation of infected mosquitoes
NOTE: This part of the protocol takes place within the BSL2 infection room. Only authorized, trained staff are permitted to enter the infection room where infected mosquitoes are housed. Mosquitoes are kept in modified cups that contain only one entry point, which automatically seals when the mouth aspirator is removed. These cups are placed inside a transparent, thermoplastic container to prevent escape. The container is located in the infection room behind a double-door system. All necessary protocols must be in place for accidental exposure to infected mosquitoes (Supplemental File S1). The protocols are country-specific and depend on the requirements of the institution.
- On days 8-10 post infection-feeding, knock the infected mosquitoes down by placing them on ice and transferring them to labeled tubes with 70% ethanol (keeping the mosquitoes of each control and treatment group separate).
- Ensure that all mosquitoes are dead before leaving the infection room.
4. Dissections of infected mosquitoes
NOTE: This part of the protocol is conducted in the laboratory.
- Transfer the mosquitoes to labeled Petri dishes lined with filter paper, keeping the control and test groups separate.
- Place a droplet of phosphate-buffered saline (PBS) on a microscope slide (marked according to the control/test group) and transfer an individual mosquito from the filter paper to the PBS.
- Remove the midgut from the immobilized, infected specimen by pinning the thorax of the mosquito with the dissecting needle whilst pulling the 7th abdominal segment with the forceps.
- With the gut being exposed and visible, look for the Malpighian tubules (Figure 4A,B) to distinguish the gut from the ovaries. Remove it from the PBS, transfer it to a droplet of 0.1% mercurochrome on a new microscope slide, and leave the gut to stain for 8-10 min.
- After staining, place a coverslip on the stained gut and view the gut under brightfield illumination at 20x-40x magnification (Figure 4C,D).
- Record the presence of and the number of oocysts per midgut for each control and treatment group (Supplemental File S2).
- Calculate the transmission-blocking activity using Equation (1):
%TBA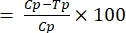 (1)
(1)
where TBA = transmission-blocking activity (reduction in oocyst prevalence); p = oocyst prevalence; C = control; and T = treatment. - Calculate the transmission-reducing activity using Equation (2):
%TRA =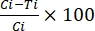 (2)
(2)
where TRA = transmission-reducing activity (reduction in oocyst intensity); I = oocyst intensity; C = control; and T = treatment.
NOTE: The TBA might not be significantly reduced, but a significant difference might be observed in the TRA and vice versa. This is dependent on the chemical material being evaluated. - Perform statistical analysis using the non-parametric t-test (Mann-Whitney).
The total number of control specimens dissected was 47, with an average to 89% prevalence and an intensity of 9.5 oocysts per midgut (Table 1, as published previously22). For the compound MMV1581558, the sample size reached a total of 42 specimens, with a 36% oocyst prevalence and an average intensity of 1.5 oocysts. This shows a reduction in oocyst prevalence of 58% and a TRA of 82% across all three biological replicates (Table 1).
Both the %TRA and %TBA for MMV1581558 were above 50%; thus, this compound could be regarded as a potential candidate for transmission-blocking and reduction. The statistical analysis for both intensity and prevalence showed a significant difference between the control group and MMV1581558 (p < 0.0001) (Table 1 and Figure 5).
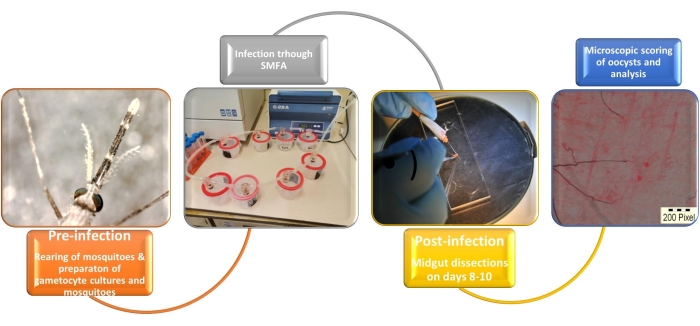
Figure 1: Illustration of the artificial infection system of Anopheles mosquitoes with Plasmodium falciparum gametocytes. Abbreviation: SMFA = standard membrane feeding assay. Please click here to view a larger version of this figure.

Figure 2: Membrane feeding system with water circulating through the glass feeders and silicone tubes to regulate the temperature of the gametocyte culture. Please click here to view a larger version of this figure.
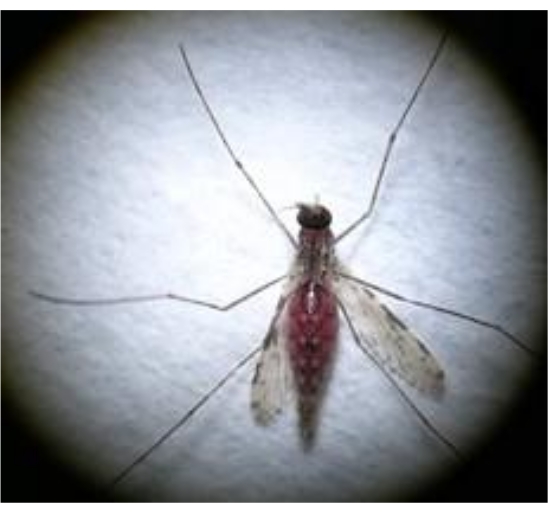
Figure 3: Fully engorged Anopheles female after a blood meal. Please click here to view a larger version of this figure.
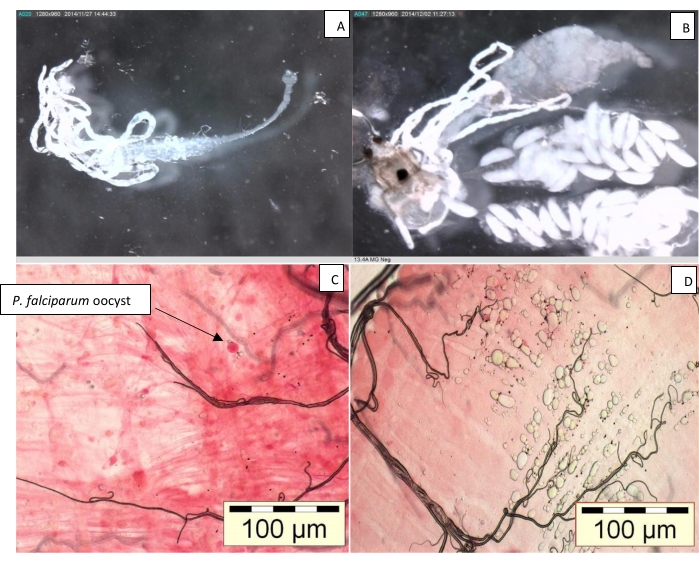
Figure 4: Midgut of Anopheles female. (A) With Malpighian tubules; (B) with ovaries; (C) with Plasmodium falciparum oocyst; and (D) uninfected midgut. Please click here to view a larger version of this figure.
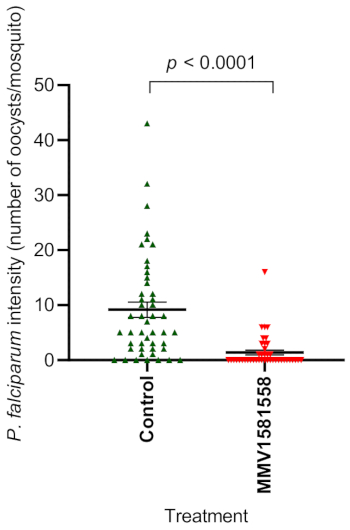
Figure 5: Statistical summary of oocyst intensity between the control group (n = 47) and the compound MMV1581558 (n = 42). Error bars, ±SE; asterisks denote significant difference (ns P > 0.05, ****P < 0.0001). Abbreviation: DMSO = dimethyl sulfoxide. Please click here to view a larger version of this figure.
| Compound | Number of reps. | Sample size | Av. Prevalence | Av. Intensity/gut | %TBA | %TRA | P-value Prevalence | P-value Intensity |
| Control | 3 | 47 | 89% | 9.5 | ||||
| MMV1581558 | 3 | 42 | 36% | 1.5 | 58% | 82% | <0.0001 | <0.0001 |
Table 1: Dataset of an antimalarial compound, MMV1581558, evaluated during three biological replicates of the standard membrane feeding assay. For each control and test group, the sample size (n) is indicated, along with the average oocyst prevalence and intensity per midgut. The transmission-blocking activity and transmission-reducing activity are indicated. Abbreviations: TBA = transmission-blocking activity; TRA = transmission-reducing activity.
Supplemental Figure S1: Infected mosquitoes contained within a chamber in the infection room of the insectary. Please click here to download this File.
Supplemental File S1: Protocol for accidental exposure to infected mosquitoes. Please click here to download this File.
Supplemental File S2: Recording sheet for oocyst prevalence and intensity. Please click here to download this File.
