Typical results from this protocol are represented below (Figure 3). As reduced cyt c absorbance locates at 550 nm, all uninhibited subsamples must show an increase in the absorbance at 550 nm. Inhibited subsamples ideally show a flat-line or slightly increasing slope (Figure 3). Slopes from inhibited subsamples are to be subtracted from uninhibited subsamples.
Samples A and B, both corrected by their correspondent inhibition and which represent NADH:cyt c oxidoreductase activity, have a similar slope (Figure 3A). However, subsamples C and D, both corrected by their correspondent inhibition and which represent succinate:cyt c oxidoreductase activity, are different, in that the activity of subsample C is higher than the activity of subsample D (Figure 3B). Note that basal absorbance can be slightly different between samples (Figure 3A).
These results (i.e., slopes already corrected by their inhibitor; Table 2) can be represented by dividing the amount of protein used (0.01 mg) as a.u./min/mg protein. From this value, the rate of cyt c reduction can be further calculated using the Lamber-Beer law21.
Importantly, these results may vary according to several factors: (i) Origin of the samples. Given that different tissues and cell types have a variable composition of OXPHOS complexes and supercomplexes, absolute values and relative changes may vary across samples. (ii) Given that different tissues may have a variable composition of OXPHOS complexes and supercomplexes, adding more frozen-thawed mitochondria (to compensate lower absolute values of a certain tissue) to the reaction mixture may have a secondary effect, which is that the ratio of Na+ or K+ per mg of protein/phospholipid in the sample decreases. Thus, caution should be taken when varying either the amount of mitochondria or the Na+/K+ concentration added to the sample. (iii) Interexperimental variation may arise from duration and temperature of freeze-thaw cycles, reagents commercial batch, or varying storage buffer of isolated mitochondria.
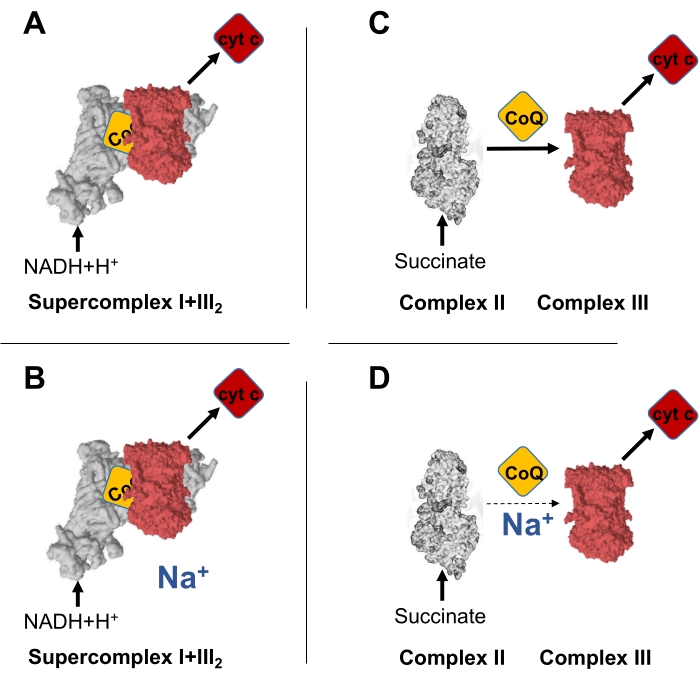
Figure 1: Na+ decreases specifically electron transfer between CII and CIII, but not between CI and CIII. (A) Schematic representation of the electron transfer between NADH and cyt c, occurring through the CoQNAD in the supercomplex I+III2. (B) Electron transfer between NADH and cyt c, occurring through the CoQNAD in the supercomplex I+III2, is not affected by intramitochondrial Na+. (C) Schematic representation of the electron transfer between succinate and cyt c, occurring through the CoQFAD in CII. (D) Electron transfer between NADH and cyt c, occurring through the CoQFAD in the supercomplex I+III2, is decreased by high intramitochondrial Na+. Please click here to view a larger version of this figure.
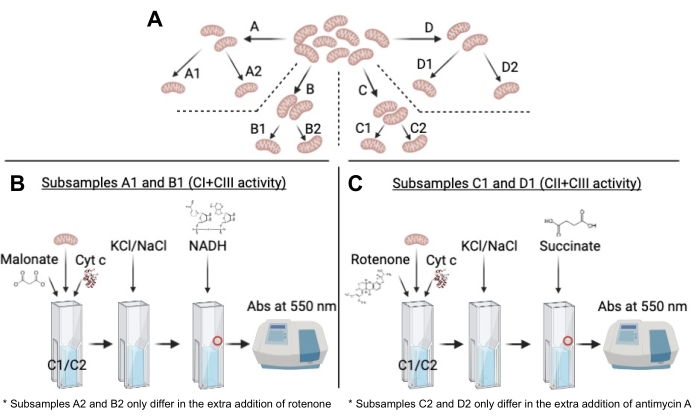
Figure 2: Schematic representation of the protocol from the subdivision of the original sample to the kinetic measurement. (A) Schematic representation of the subsample division, highlighting the same origin of all subsamples. (B) Scheme of the successive steps of reagent's additions for CI+CIII activity in subsamples A1 and B1. The red circle represents the location where NADH should ideally be added. Note that the only difference with subsamples A2 and B2 is the extra addition of rotenone in the latter. (C) Scheme of the successive steps of reagent's additions for CII+CIII activity in subsamples C1 and D1. The red circle represents the location where succinate should ideally be added. Note that the only difference with subsamples C2 and D2 is the extra addition of antimycin A in the latter. Please click here to view a larger version of this figure.
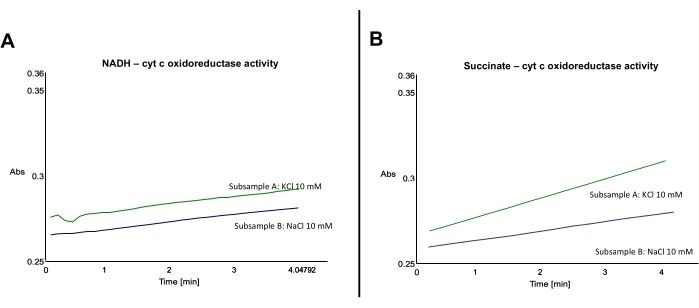
Figure 3: Effect of Na+ on cyt c reduction by mouse liver mitochondrial membranes upon NADH or succinate addition. (A) Representative traces showing the effect of Na+ on cyt c reduction by mouse liver mitochondrial membranes oxidizing NADH. (B) Representative traces showing the effect of Na+ on cyt c reduction by mouse liver mitochondrial membranes oxidizing succinate. Please click here to view a larger version of this figure.
| Compound | Concentration |
| K2HPO4 | 25 mM |
| MgCl2 | 5 mM |
| KCN | 3 mM |
| Bovine Serum Albumin (BSA) | 2.5 mg/mL |
Table 1: Composition of C1/C2 buffer. Buffer composition is presented in molar concentrations.
| Expected rates | +KCl (Mean) | +KCl (SD) | +NaCl (Mean) | +NaCl (SD) | Mann-Whitney P value |
| CII + CIII (n = 4) | 0.050659 | 0.0068377 | 0.023217 | 0.0024511 | 0.0286 |
| Individual values | 0.0509629 | 0.02250151 | |||
| 0.0561086 | 0.02664035 | ||||
| 0.0393956 | 0.01984683 | ||||
| 0.0561695 | 0.0238827 | ||||
| CI + CIII (n = 4) | 0.016681 | 0.00237326 | 0.017756 | 0.0029472 | 0.4857 |
| Individual values | 0.01610133 | 0.01780299 | |||
| 0.01878711 | 0.01901848 | ||||
| 0.01303777 | 0.01308397 | ||||
| 0.01879871 | 0.02112066 | ||||
Table 2: Expected rates ranges. The expected values for each activity are presented in arbitrary units. The corresponding statistical test between +KCl and +NaCl is also presented. "n" represents the number of replicates.
