Over the last century, X-ray crystallography has been critical in elucidating and understanding the structure-function paradigm of biological macromolecules. To date, it continues to be one of the most successful methods in elucidating atomic resolution structures of many uniquely different proteins that are crucial to the fundamental understanding of cell biochemistry, medicine, and early drug discovery1,2. However, protein crystallization remains a bottleneck in studying many protein targets, particularly membrane proteins and large protein complexes3. Consequently, protein crystallization is almost always considered an art due to the labor-intensive trial-and-error approaches employed4,5,6.
A precipitating agent is usually added to a protein solution at high concentration to form a well-ordered, regular, and repeating lattice arrangement of protein molecules, known as crystals. Under favorable conditions, such as temperature, pH, concentration, and precipitant agent, a supersaturated solution eventually forms, followed by crystal nucleation and growth7,8. Although there have been many advances in crystallization trial setups, predominantly with the development of high-throughput robotic systems and the availability of ready-made "sparse matrix" screens, the general approaches to protein crystallization have largely remained unchanged over the years. Common experimental protein crystallization techniques include vapor diffusion (hanging drop and sitting drop)9, microbatch (under oil)10,11, free-interface diffusion (microfluidic devices)12, and dialysis (using buttons and other techniques)13,14,15. However, other more specialized setups also exist, such as mesophase approaches for crystallizing membrane proteins16,17. While the majority of X-ray protein structures deposited in the Protein Data Bank have so far been solved through crystallization by vapor diffusion methods6,18,other approaches, such as crystallization by dialysis, seem to be underutilized, likely due to the practical aspects related to their experimental setup.
Crystallization by dialysis simply relies on the slow diffusion of solutes (precipitants, ions, additives, and buffers) through a semi-permeable membrane that simultaneously prevents protein molecules from circulating. This way, the protein solution is slowly brought into equilibrium, with the precipitant reaching the necessary concentration to crystallize. The system's kinetics depends on temperature, precipitant concentration, and the cellulose membrane molecular weight cut-off (MWCO)19. To date, the most popular crystallization setup by dialysis has been using microdialysis buttons made of transparent acrylic sheets. These are usually immersed in reservoirs (mostly using vapor diffusion hanging drop plates) containing the crystallization precipitant solutions. However, this lower-throughput method also requires specific assembly to seal the protein solution within the dialysis membrane placed over the button chamber, as illustrated in Figure 1. Moreover, air bubbles trapped between the dialysis membrane and the protein solution are a frequent problem that impair crystal growth. Another constraint of the method is the sample requirements, whereby much higher concentrations and volumes are necessary compared to vapor diffusion methods, to accommodate the dialysis buttons. Therefore, crystallization using microdialysis buttons has been perceived as an unappealing method, especially for difficult targets such as membrane proteins, whose purification yields are frustratingly low. Recently, microfluidic devices have been developed to facilitate protein crystallization by dialysis15. These chips have also been designed to have high X-ray transparency with low background, allowing the chips to be used for in situ data collection at room temperature, thus eliminating the inconvenience of harvesting and cryocooling crystals. Despite these advances, the approach is still very low-throughput and expensive.
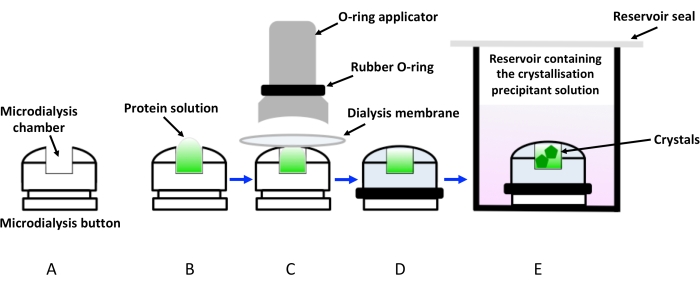
Figure 1: Schematic representation of crystallization by dialysis using dialysis buttons. (A) Schematic representation of a crystallization dialysis button. (B) The protein solution is added to the microdialysis button chamber. (C) The dialysis membrane is held to the microdialysis button with the help of a rubber ring (O-ring) applied via an applicator. (D) The dialysis button is ready to be immersed in the reservoir containing the crystallization solution (dialysis solution), as shown in (E). The vial containing the immersed dialysis button must be sealed to avoid evaporation. Please click here to view a larger version of this figure.
Here, a straightforward protocol is presented for screening protein crystallization conditions and crystal growth using the 96-well high-throughput dialysis plate. These disposable plates are designed to be used similarly to the vapor diffusion crystallization plates (pipette then seal), as shown in Figure 2. The plates can accommodate up to 3.2 µL of protein and 350 µL of dialysis solution. Each well features a separate regenerated cellulose membrane to prevent cross-contamination between the wells. The setup takes around 10 min to complete and does not require any specialized equipment besides what can be found in all standard crystallization laboratories. Four different proteins, including two membrane proteins, are used to demonstrate and validate this approach as an effective method for high-throughput (HTP) protein crystallography.
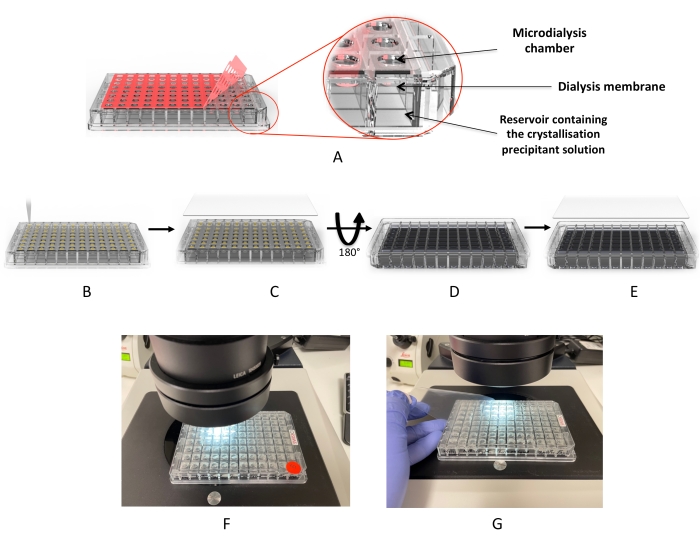
Figure 2: Crystallization workflow using the microdialysis plate. (A) Removal of the red adhesive "cover film". (B) Dispensing the protein droplets into each of the drop wells. (C) The wells are covered with the UV "cover film". (D) The plate is inverted to add the dialysis solutions (or crystallization screen). (E) The plate is sealed and incubated. (F,G) Microscope inspection of the drops. Please click here to view a larger version of this figure.
The use of this crystallization by dialysis protocol was demonstrated using the 0.5 mL dialyzer tube (Figure 3) for the large-scale (hundreds to thousands) production of microcrystals, suitable for state-of-the-art data collection methods such as serial crystallography at both XFEL facilities20,21,22,23,24 and synchrotrons25,26,27, as well as for MicroED28,29,30 approaches.
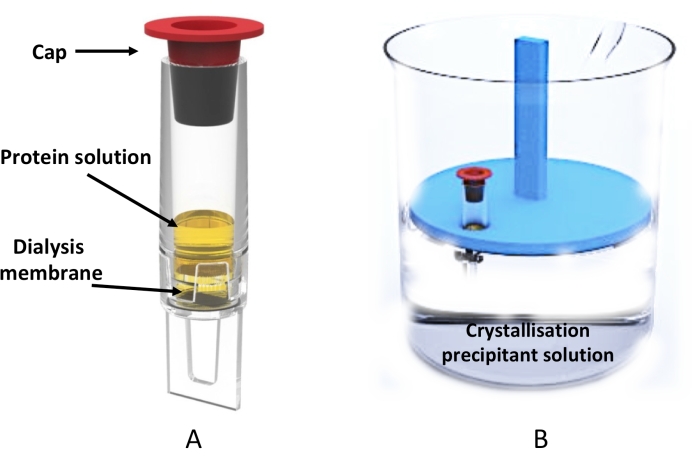
Figure 3: Large-scale microdialysis crystallization using the dialyzer tube. (A) Schematic representation of the 0.5 mL dialyzer tube. (B) Side view of a beaker containing the crystallization solution and the floating tube rack holding a dialyzer tube. Please click here to view a larger version of this figure.
Four proteins were crystallized using the microdialysis plate (with 10 kDa MWCO membranes), including two membrane proteins. The chicken egg-white lysozyme from the lyophilized powder (see Table of Materials) was prepared at 50 mg.mL-1 in 20 mM NaOAc (pH 4.5), and the lyophilized thaumatin (see Table of Materials) was dissolved in water to a final concentration of 25 mg.mL-1. The two membrane proteins used in this study were the E. coli multidrug efflux pump AcrB and the E. coli lactose transporter LacY. AcrB was expressed using C43 (DE3) cells and purified by Ni-NTA affinity chromatography and size-exclusion chromatography in 10 mM Tris (pH 7.5), 300 mM NaCl, 0.03% (w/v) n-dodecyl-β-D-maltoside (DDM), and 5% (v/v) glycerol32. Subsequently, the protein was concentrated using a 100 kDa MWCO centrifugal concentrator to a final concentration of 6 mg.mL-1. LacY was also expressed in C43 (DE3) cells, purified by Ni-sepharose affinity chromatography followed by size-exclusion chromatography in 20 mM Tris (pH 7.5), 150 mM NaCl, 0.03% (w/v) DDM, and concentrated using a 100 kDa MWCO concentrator to 10 mg.mL-1 4,33.
For the lysozyme and thaumatin proteins, the 96-well microdialysis plate was filled with a crystallization grid screen prepared in-house as the dialysis solution, while for the membrane protein AcrB, a grid screen was prepared in-house from a previously published crystallization condition34. For LacY, initial hit conditions were found from a commercial screen (see Table of Materials) and further optimized with a grid screen made in-house. The crystallization drop ratio for lysozyme, thaumatin, and AcrB was 1:100, with 2 µL of protein to 200 µL of precipitant (dialysis solution). LacY crystallization drops were also at a 1:100 ratio, with 1 µL of protein to 100 µL of dialysis solution, due to a lower amount of available protein.
Crystals from all four proteins, including the membrane proteins, started to appear between 1-8 h at 20 °C following setup with the microdialysis plate. In this case, it was not necessary to supplement the dialysis solution with detergent for the membrane protein crystallization, typically done during the standard dialysis process to prevent membrane protein aggregation, as the detergent micelles were expected to be larger than the MWCO of the dialysis membranes35.
Following the successful crystallization of the proteins on the microdialysis plate (Figure 4), crystallization conditions were noted, and large-scale crystallization by dialysis was performed using the dialyzer tube with the same 10 kDa MWCO dialysis membranes. Thousands of microcrystals for each of the individual proteins grew inside the dialyzer, as shown in Figure 5. For thaumatin crystallization, 250 µL of protein was loaded onto the dialyzer and dialyzed against 50 mL (1:200 ratio). For AcrB, 250 µL of protein was dialyzed against 25 mL of dialysis solution (1:100). LacY crystallization was also set up at the same ratio with 100 µL of protein to 10 mL of dialysis solution.
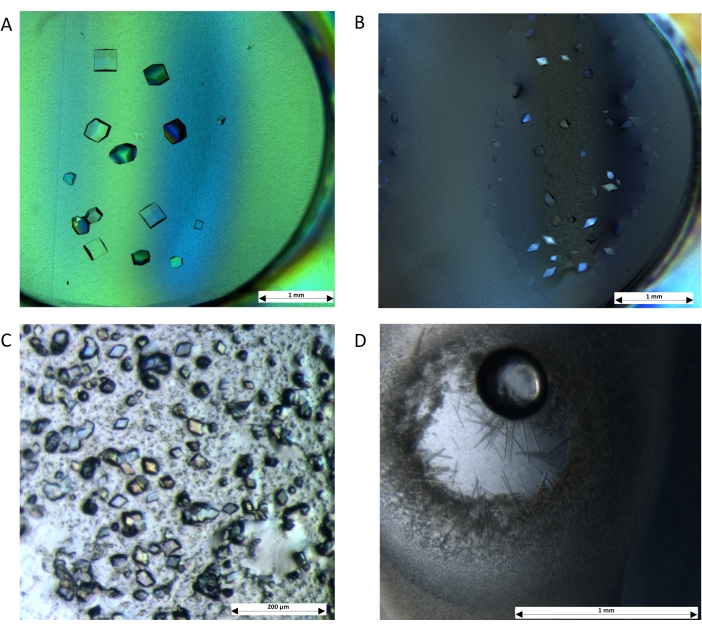
Figure 4: Crystals are grown by dialysis using the microdialysis plate. The protocol demonstrates the applicability of the setup with soluble proteins: (A) Lysozyme was dialyzed against 0.1 M NaOAc (pH 4.0), 0.5 M NaCl, and 25% (v/v) glycerol. (B) Thaumatin was dialyzed against 0.1 M Bis-tris propane (pH 6.6), 1 M K/Na tartrate, and 18% (v/v) ethylene glycol. Crystals were also obtained for membrane proteins: (C) AcrB was dialysed against 0.1 M MES (pH 5.5), 0.3 M NaCl, and 20% (v/v) PEG-400. (D) LacY was dialysed against 0.1 M MES (pH 6.5), 0.1 M NaCl, and 32% (v/v) PEG-300. Images were captured using a stereo high-magnification microscope with a cross polariser. Scale bars: (A,B,D) = 1 mm; (C) = 200 µm. Please click here to view a larger version of this figure.

Figure 5: Representative images of membrane protein crystals grown by dialysis using the dialyzer tube. (A) Image showing the tube full of microcrystals (indicated by the black arrow). (B) Microscopic image of 2 µL from a slurry of thaumatin crystals grown by dialysis using the 0.5 mL dialyzer tube, dialyzed against 0.1 M Bis-Tris propane (pH 6.6), 1.4 M K/Na tartrate, and 18% (v/v) ethylene glycol. (C) Dark-field (left) and bright-field (right) images of AcrB microcrystals grown by dialysis against 0.1 M MES (pH 5.5), 0.3 M NaCl, and 20% (v/v) PEG-400. (D) Dark-field (left) and bright-field (right) images of LacY microcrystals grown by dialysis against 0.1 M MES (pH 6.5), 0.1 M NaCl, and 32% (v/v) PEG-300. Some precipitates are observed. Scale bars: (B,D) = 200 µm; (C) = 500 µm. Please click here to view a larger version of this figure.
