Effects of crocetin on cell viability
Crocetin at 0.1 µM, 0.5 µM, 1 µM, 5 µM, 10 µM, 50 µM, and 100 µM had a significant proliferative effect on cells, while crocetin at concentrations above 200 µM significantly inhibited the proliferation of H9c2 cells (Figure 1A). After 4 h of treatment with 400 µM H2O2, the cell viability was reduced considerably, and crocetin could reverse this change to a certain extent (Figure 1B). Since no significant difference between 10 µM and 100 µM crocetin was observed on the H2O2-induced H9c2 cell viability, 10 µM crocetin was chosen as the high concentration, and 1 µM and 0.1 µM were used as the medium and low dose groups, respectively.
Effects of crocetin on LDH, CK, MDA, SOD, GSH-Px, and CAT in H9c2 cells
After 4 h of treatment with 400 µM H2O2, the levels of LDH, CK, and MDA increased appreciably, while the levels of SOD, GSH-Px, and CAT decreased. Pretreatment of 10 µM crocetin for 24 h can reverse the above changes and shows an obvious dose-dependent effect. As a positive control drug, coenzyme Q10 can only change the levels of CK, MDA, and SOD (Figure 2).
The effect of crocetin on ROS in H9c2 cardiomyocytes
As a blank control, H9c2 cardiomyocytes expressed almost no ROS. At the same time, 400 µM H2O2 for 4 h treatment could enhance the ROS level notably, which can be reversed by 10 µM crocetin to some extent (Figure 3). The fluorescence results showed that green fluorescence was very weak in the normal group. In comparison, 400 µM H2O2 for 4 h treatment could enhance the green fluorescence signal, and this enhancement could be reduced by 10 µM crocetin (Figure 3).
Effects of crocetin on H2O2-induced mitochondrial membrane potential and apoptosis
JC-1 staining showed more red fluorescence and less green fluorescence in the blank control group. After 4 h of treatment of 400 µM H2O2, more green fluorescence and less red fluorescence were observed, and 10 µM crocetin could reverse this change to some extent (Figure 4A). TUNEL staining results showed that apoptosis-related signaling was not detected in the blank control group, while the apoptosis-related signaling was appreciably enhanced after 400 µM H2O2 for 4 h of treatment, which could be reversed by 10 µM crocetin to some extent (Figure 4B).
Effects of crocetin on H2O2-induced excessive autophagy
H9c2 cardiomyocytes in the blank control group showed no obvious autophagy flow. Fluorescence showed the appearance of punctate yellow spots in H9c2 cardiomyocytes pretreated with 400 µM H2O2 for 4 h, indicating an obvious over-activation of autophagy. However, this change was reversed after the 10 µM crocetin pretreatment. In the control group, the Ad-mCherry GFP-LC3B virus could only be observed as a weak diffuse yellow background by fluorescence. However, punctate yellow spots were observed after 400 µM H2O2 for 4 h of treatment, and this change was reversed after the 10 µM crocetin pretreatment (Figure 5).
Detection of crocetin on the expression of mitophagy-related proteins
Western blot results showed that in the control group, the expression levels of PINK1 and Parkin were lower. In 4 h H2O2-stimulated H9c2 cardiomyocytes, the expression levels of PINK1 and Parkin increased, while the 10 µM crocetin pretreatment could reduce the increase of PINK1 and Parkin (Figure 6).
Detection of the effect of crocetin on the translocation of Parkin mitochondria
The immunofluorescence results showed that the red fluorescence signal representing Parkin in the blank control group was very weak; however, after 400 µM H2O2 for 4 h of treatment, the red fluorescence signal was enhanced, and colocalization with the green fluorescence representing Tom20 increased. After the pretreatment of 10 µM crocetin, the red fluorescence signal was weakened, and colocalization with the green fluorescence signal was reduced (Figure 7).
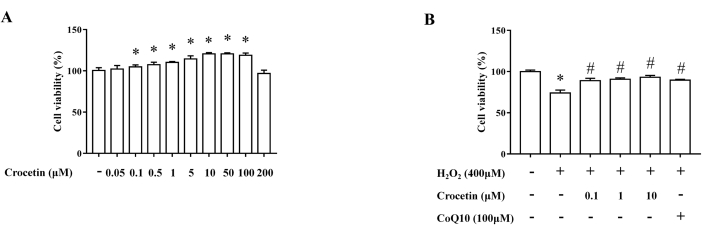
Figure 1: Detection of cell viability by the MTT assay. (A) Effects of crocetin at different concentrations on cell viability (n = 6). (B) Effects of crocetin at different concentrations on cell viability after H202 intervention (n = 6). *p < 0.05 versus control group, #p < 0.05 versus H2O2 treatment group. Please click here to view a larger version of this figure.
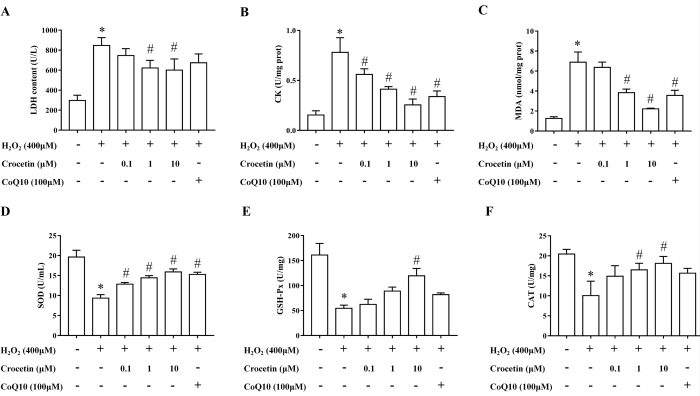
Figure 2: Detection of LDH, CK, MDA, SOD, GSH-Px, and CAT levels. (A) LDH level in cell supernatant (n = 6). (B) CK level in cell lysate (n = 6). (C) MDA level in cell lysate (n = 6). (D) SOD level in cell supernatant (n = 6). (E) GSH-Px level in cell lysate (n = 6). (F) CAT level in cell lysate (n = 6). *p < 0.05 versus blank control group, #p < 0.05 versus H2O2 treatment group. Please click here to view a larger version of this figure.
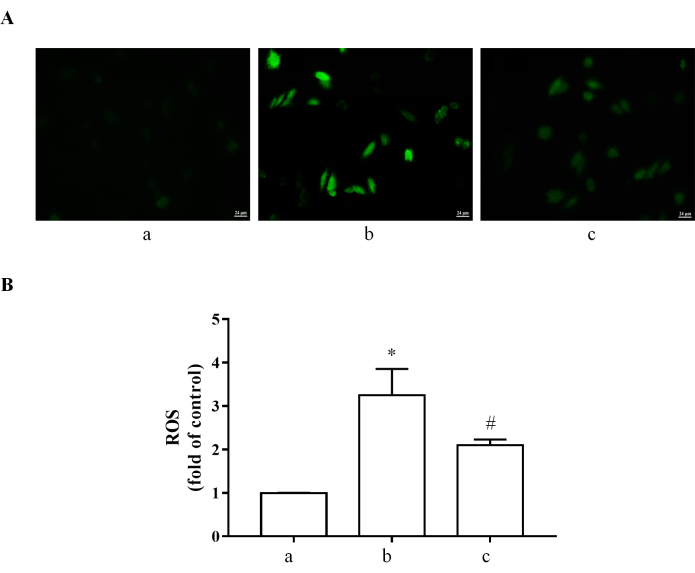
Figure 3: ROS determined by DCFH-DA. (A) DCFH-DA was used to measure ROS levels in H9c2 cardiomyocytes. ROS: green. (B) Quantification data of ROS. (a) H9c2 cardiomyocytes without treatment. (b) H9c2 cardiomyocytes stimulated with 400 µM H2O2 for 4 h. (c) H9c2 cells pretreated with 10 µM crocetin for 24 h and then stimulated with 400 µM H2O2 for 4 h. Scale bar = 24 µm. *p < 0.05 versus blank control group, #p < 0.05 versus H2O2 treatment group. Please click here to view a larger version of this figure.
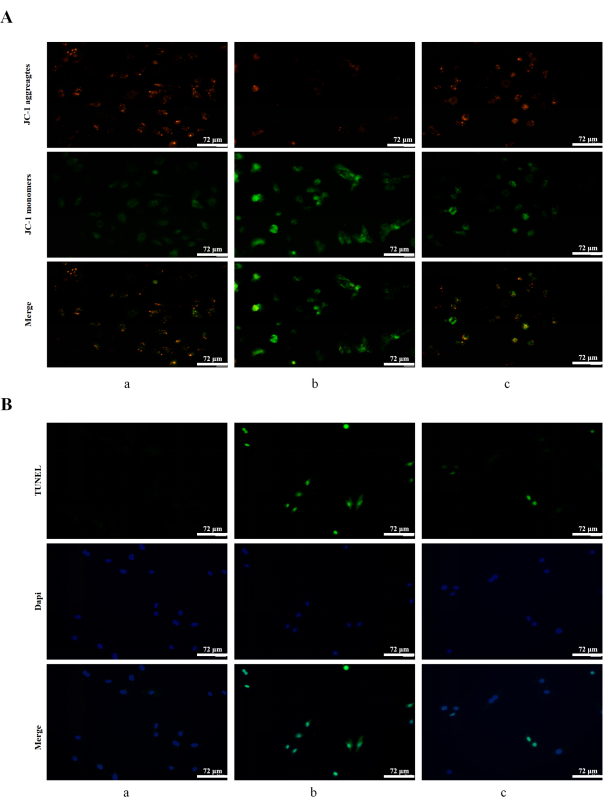
Figure 4: Detection of mitochondrial membrane potential and apoptosis. (A) Mitochondrial membrane potential was determined by JC-1 staining. JC-1 aggregates: red; JC-1 monomers: green. (B) Apoptosis was detected by TUNEL staining in H9c2 cells. TUNEL: green; DAPI: blue. (a) H9c2 cardiomyocytes without treatment. (b) H9c2 cardiomyocytes stimulated with 400 µM H2O2 for 4 h. (c) H9c2 cells pretreated with 10 µM crocetin for 24 h and then stimulated with 400 µM H2O2 for 4 h. Scale bars = 72 µm. Please click here to view a larger version of this figure.
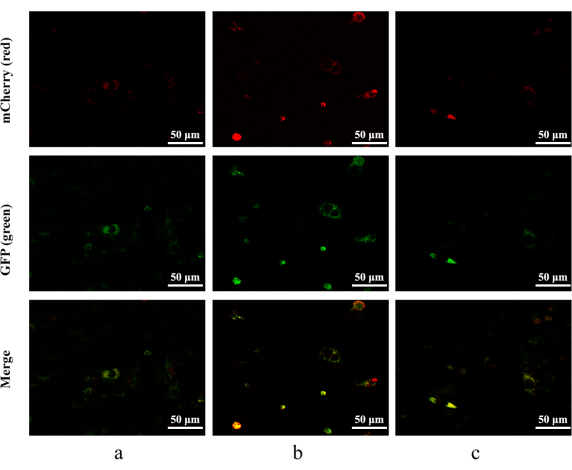
Figure 5: Autophagic flux detected by mCherry-GFP-LC3B adenovirus. mCherry: red; GFP: green. (a) H9c2 cardiomyocytes without treatment. (b) H9c2 cardiomyocytes stimulated with 400 µM H2O2 for 4 h. (c) H9c2 cells pretreated with 10 µM crocetin for 24 h and then stimulated with 400 µM H2O2 for 4 h. Scale bars = 50 µm. Please click here to view a larger version of this figure.
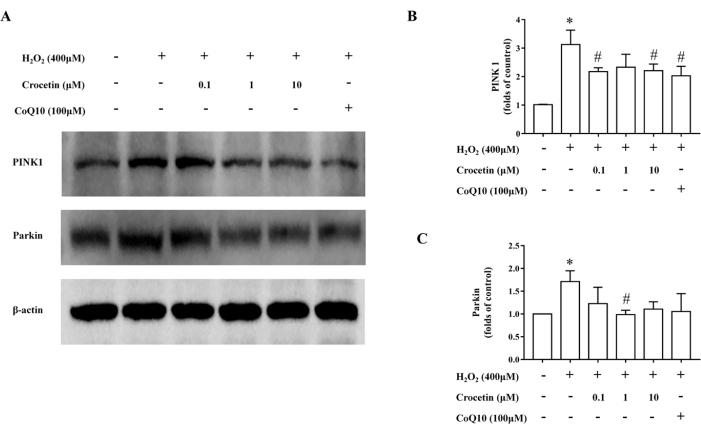
Figure 6: The content of mitophagy-related proteins was detected by western blotting. (A) Representative western blot illustrating PINK1 and Parkin expression. β-actin was adopted as an internal reference. (B) Relative PINK1 expression (n = 3). (C) Relative Parkin expression (n = 3). *p < 0.05 versus control group, #p < 0.05 versus H2O2 treatment group. Please click here to view a larger version of this figure.
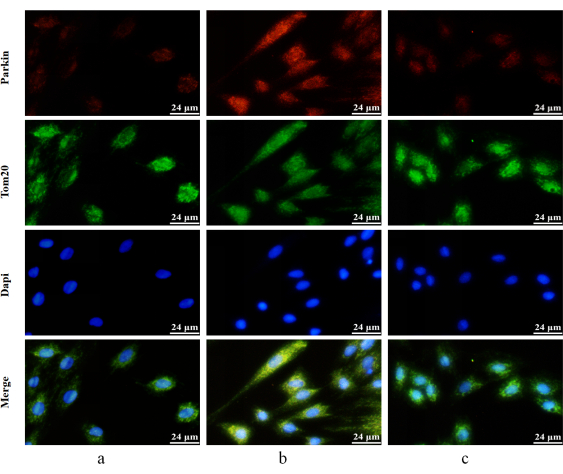
Figure 7: Detection of Parkin's mitochondrial translocation by immunofluorescence double staining. Red fluorescence-labeled Parkin's protein and green fluorescence labeled-Tom20 protein. Parkin: red; Tom20: green; DAPI: blue). (a) H9c2 cardiomyocytes without treatment. (b) H9c2 cardiomyocytes stimulated with 400 µM H2O2 for 4 h. (c) H9c2 cells pretreated with 10 µM crocetin for 24 h and then stimulated with 400 µM H2O2 for 4 h. Scale bars = 24 µm. Please click here to view a larger version of this figure.
Supplementary File 1: The working instructions of LDH, CK, MDA, SOD, GSH-Px, and CAT assays. Please click here to download this File.
