Dual Immunofluorescence of γH2AX and 53BP1 in Human Peripheral Lymphocytes
Summary
This protocol presents a method to assess the formation and repair of DNA double-strand breaks through the simultaneous detection of γH2AX and 53BP1 foci in interphase nuclei of bleomycin-treated human peripheral lymphocytes.
Abstract
Double strand breaks (DSBs) are one of the most severe lesions that can occur in cell nuclei, and, if not repaired, they can lead to severe outcomes, including cancer. The cell is, therefore, provided with complex mechanisms to repair DSBs, and these pathways involve histone H2AX in its phosphorylated form at Ser-139 (namely γH2AX) and p53 binding protein 1 (53BP1). As both proteins can form foci at the sites of DSBs, identification of these markers is considered a suitable method to study both DSBs and their kinetics of repair. According to the molecular processes that lead to the formation of γH2AX and 53BP1 foci, it could be more useful to investigate their co-localization near the DSBs in order to set up an alternative approach that allows quantifying DSBs by the simultaneous detection of two DNA damage markers. Thus, this protocol aims to assess the genomic damage induced in human lymphocytes by the radiomimetic agent bleomycin through the presence of γH2AX and 53BP1 foci in a dual immunofluorescence. Using this methodology, we also delineated the variation in the number of γH2AX and 53BP1 foci over time, as a preliminary attempt to study the repair kinetics of bleomycin-induced DSBs.
Introduction
DNA damage is continuously induced by agents that can be endogenous, such as ROS generated by cellular oxidative metabolism, or exogenous, both chemicals and physical1. Among the most harmful lesions, double-strand breaks (DSBs) play a fundamental role in contributing to genomic instability, since they cause chromosome aberrations that in turn can initiate the carcinogenesis process. Thus, cells are provided with complex and efficient mechanisms of DSBs repairing2.
When a DSB occurs, the cell triggers the DNA damage response (DDR) where, together with the MRE11/RAD50/NBS1 complex, ATM or ATR kinases are recruited to activate other proteins that slow down or stop the cell cycle3. An essential target of these kinases is histone H2AX, which is phosphorylated on Ser-139 within a few megabases from the DSBs (namely γH2AX), thereby allowing the recruitment of several repair factors such as, among others, BRCA1 and p53 binding protein 1 (53BP1)3. Later, one pathway among homologous recombination (HR), non-homologous end joining (NHEJ), or single-strand annealing (SSA) is triggered to repair the DSBs4,5. Therefore, 53BP1 is involved in dictating the choice between HR or NHEJ, mainly promoting the activation of NHEJ rather than HR6. Moreover, both the phosphorylated form of H2AX histone and 53BP1 can form foci at the sites of DSBs. As these foci persist until the integrity of the double strand is restored, assessing the appearance/disappearance of γH2AX or 53BP1 foci within a time interval is considered a useful method to evaluate the occurrence and repair of DSBs in a cell system6,7. However, according to the molecular processes above described, since γH2AX and 53BP1 foci are expected to co-localize near the DSBs during DDR8,9, it can be useful to detect concurrently the presence of these markers in a dual immunofluorescence.
Thus, the aim of this manuscript was to evaluate the suitability of the simultaneous quantification of γH2AX and 53BP1 foci to assess the genomic damage induced in human peripheral lymphocytes by the radiomimetic agent bleomycin. Using the same methodology, we also attempted to delineate repair kinetics of bleomycin-induced DSBs according to a previously set up experimental procedure10.
Protocol
The study was approved by the ethical committee of Pisa University, and informed and signed consent was obtained from each donor.
1. Formation of γH2AX and 53BP1 foci
- Preparation of samples and mutagenic treatment
- Collect whole blood samples by venipuncture from healthy adult individuals in blood collection (e.g., Vacutainer) tubes containing lithium heparin as an anticoagulant.
- In order to guarantee proper blood sample preservation, start the procedure within 24 h of sampling.
- Add 300 µL of the sample to a tube containing 4.7 mL of complete medium (0.5% penicillin-streptomycin, 0.75% phytohemagglutinin, 10% FBS previously inactivated, 88.75% RPMI 1640).
- Then add bleomycin sulfate to a final concentration of 5 µg/mL.
CAUTION: Bleomycin sulfate is a mutagen. Avoid skin contact and inhalation. Prepare the solution and add the sample under a hood. - For each sample, set up a negative control (absence of mutagen).
- Place the tube in a thermostat at 37 °C for 2 h.
- Fixation
- Centrifuge samples at 540 x g for 5 min at room temperature.
- Aspirate the supernatant and resuspend the pellet with vortex.
- Add 5 mL of hypotonic solution (2.87 g of KCl dissolved in 500 mL of deionized water) and 400 µL of pre-fixative solution (5:3 acetic acid: methanol) to cause hemolysis of red blood cells.
- Centrifuge samples at 540 x g for 5 min at room temperature.
- Aspirate the supernatant and resuspend the pellet in 5 mL of methanol at room temperature for at least 30 min to fix the cells.
- Alternatively, store the cells at -20 °C until use.
- Slides preparation
- Centrifuge samples at 540 x g for 5 min at room temperature.
- Aspirate the supernatant and resuspend the pellet in 5 mL of a 3:1 methanol: acetic acid solution. Repeat these steps one more time.
- At the end, centrifuge the solution again at 540 x g for 5 min at room temperature.
- Aspirate the supernatant again leaving enough solution (0.5 mL) to resuspend the pellet, pipette vigorously, drop the resuspended cell pellet on the slides, and air dry. Store the slides at 4 °C.
- Immunofluorescence
NOTE: Immunofluorescence is an immunologic method for identifying specific cell targets using a primary antibody binding the target itself and a fluorescent secondary antibody binding the primary that allows localizing the target11. In this case, a mouse monoclonal anti-53BP1 (1:50) and a rabbit polyclonal anti-γH2AX (1:50) are used as primary antibodies, while AlexaFluor568 anti-mouse (1:400) and DyLight488 anti-rabbit (1:200) are used as secondary antibodies, respectively.- Wash the slides two times in 50 mL of 1x PBS for 5 min in couplin jar (16 slides back-to-back).
- Then keep them for 30 min in blocking solution (10 mL of FBS, 10 mL of 10x PBS, 80 mL of deionized water, 300 µL of Triton-X).
- Add to each slide 10 µL of each of the two solutions containing the primary antibodies dissolved in the blocking solution. Cover the slides with paraffin tape and incubate at 4 °C overnight.
- After incubation, perform three washing in 1x PBS for 5 min.
- Add to each slide 10 µL of each of the two solutions containing the secondary antibodies dissolved in the blocking solution. Cover the slides with paraffin tape and incubate at room temperature for 2 h.
- Perform three washing in 1x PBS for 5 min.
- Add 2.5 µL of antifade solution with DAPI on the coverslips before the assembly to counterstain the nuclei.
NOTE: In order to assess foci kinetics, the procedure is the same as described from 1.1 to 1.4, noting that cell harvesting and the dual immunofluorescence are performed at 0 h, after 2 h post-bleomycin treatment and then, after having removed the mutagen, at 4 h, 6 h, and 24 h post-treatment.
2. Analysis through a fluorescence microscope
NOTE: "Fluorescence microscope" refers to any microscope that uses fluorescence to generate an image. The specimen is illuminated with light of a specific wavelength (or wavelengths) which is absorbed by the fluorophores, causing them to emit light of longer wavelengths12. AlexaFluor568 and DyLight 488 absorb light of approximately 568 and 488 nm and emit light of 603 and 520 nm, respectively. Thus, they are visible as red or green fluorescence using a TRITC or a FITC filter, respectively.
- Score the presence of foci in each slide under a 100x immersion objective (Figure 1).
- Score 200 nuclei per slide and count the number of γH2AX/53BP1 foci in each nucleus.
- Express results in terms of the average number of foci per total nuclei scored. These include both γH2AX/53BP1 positive (showing at least one fluorescence signal) and negative (not showing any fluorescence signal) nuclei.
Representative Results
Data obtained by the fluorescence microscope analysis of peripheral lymphocytes allow us to evaluate three main aspects: the effectiveness of bleomycin treatment in increasing the number of γH2AX and 53BP1 foci (and thus of DSBs) due to its mutagenic effect, at what extent both foci co-localized at the site of DSBs, and the time-course of γH2AX and 53BP1 foci to delineate the repair kinetics of bleomycin-induced DSBs. As expected, a very higher frequency of both γH2AX and 53BP1 foci was observed between untreated and treated cells, thus confirming that bleomycin induces the formation of DSBs in peripheral lymphocytes (Figure 2).
A large difference was observed in the number of foci of the two markers; in particular, γH2AX foci were more numerous than 53BP1 foci, thus indicating that co-localization does not always occur and can depend on multiple factors (Figure 3).
Regarding the repair kinetics of DSBs, first of all, it is important to underline how the actual repair of foci can start before the first selected time point, which is 2 h, and that at various time points, we can observe newly formed foci as well as foci that have been formed previously and that have not been repaired yet.
Taking into account what was previously said, the time-course of γH2AX and 53BP1 foci over time showed a different behavior although they contribute to the same function. γH2AX foci increased after 2h post-treatment, and then they started a progressive reduction, without, however, reaching the control value at 24 h after the treatment. At variance, the frequency of 53BP1 foci increased until 4 h post-treatment, then decreased at 6 h and returned to the highest value 24 h post-treatment (Figure 4).
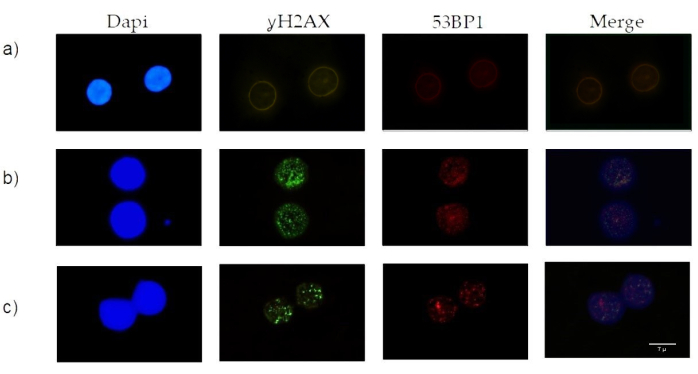
Figure 1: Dual immunofluorescence in human peripheral lymphocytes for γH2AX and 53BP1: (A) nuclei without γH2AX and 53BP1 foci before bleomycin treatment, (B) and (C) nuclei with different amounts of γH2AX and 53BP1 foci due to bleomycin treatment. Please click here to view a larger version of this figure.
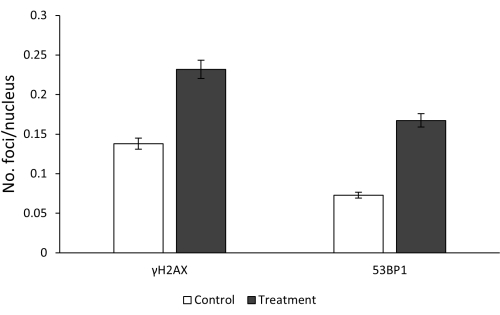
Figure 2: Comparison between untreated and bleomycin-treated (5 µg/mL) lymphocytes for the two DSBs markers. Error bars represent SEM. Please click here to view a larger version of this figure.
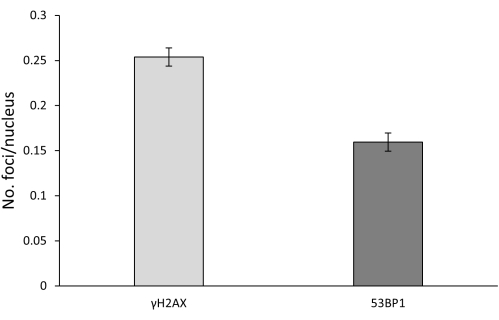
Figure 3: Comparison between the total amount (both spontaneous and mutagen-induced) of γH2AX and 53BP1 foci. Error bars represent SEM. Please click here to view a larger version of this figure.
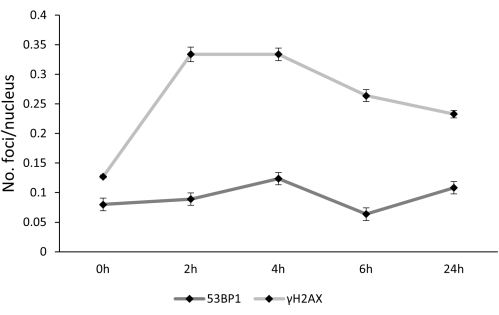
Figure 4: γH2AX and 53BP1 foci kinetics. 0 h, 2 h, 4h, 6 h, and 24 h represent the different times at which lymphocytes were harvested; error bars represent SEM. Please click here to view a larger version of this figure.
Discussion
Immunofluorescence analysis of γH2AX and 53BP1 foci is a suitable method for assessing genomic damage in interphase nuclei of a cell system. This procedure has several critical points that can affect the outcome of the experiments, mainly, the agents used in fixation and permeabilization, the type of antibodies and their dilution factors, and the concentration of the mutagen.
The maintenance of protein integrity is fundamental since the immunofluorescence method expects to identify antigens that are mainly proteins. In this protocol, methanol is used to fix lymphocytes. This step is particularly important since alcohol acts by displacing water and causing the precipitation of soluble proteins, while the other not fixed cellular components can be removed by washing in a saline buffer solution, such as PBS13.
Another key passage is permeabilization which is performed through a blocking solution, which in this case allows both permeabilization and blocking. Blocking is performed by adding FBS to the solution: it binds the proteins in the sample and prevents antibodies from binding the wrong target since the bond between FBS and the specific target of the antibody can later be broken due to the high affinity between the antibody and the target itself. Permeabilization can be performed using several solvents, in this case, the nonionic detergent Triton X-100 is used: it intercalates into a phospholipid bilayer, solubilizing the plasma membrane and then disrupting it14.
As the choice of efficient γH2AX and 53BP1 antibodies is crucial for the success of the immunofluorescence, the use of antibodies of proven reliability is strongly recommended. It is also suggested to try several dilutions of the antibodies to identify the best-performing concentration.
As preliminary experiments, we studied different controls in order to assess possible background noise or autofluorescence. Notably, we used, in different experiments, the primary antibodies only, the secondary antibodies only, each stain individually, and no staining. As expected, in the presence of primary/secondary antibodies only, we did not observe any focus, while we observed either green or red fluorescent spots (indicating the presence of either γH2AX or 53BP1 foci) when we performed the protocol with each stain individually. With regard to no staining controls, we observed background noise/autofluorescence, which manifests itself with diffuse staining. Instead, after performing the immunofluorescence protocol, γH2AX or 53BP1 foci are clearly visible in the brown-dark nuclei as green or red signals under the appropriate filter for the used fluorochromes (FITC or TRITC). Thus, background noise/autofluorescence does not influence in a significant way the identification of foci, also because we used specific criteria in order to assess if one fluorescent spot is a proper focus (i.e., during the focusing of the microscope, γ-H2AX and 53BP1 foci should not be on the same focal plane of other fluorescent spots potentially present outside the cell nucleus). These criteria have been chosen since they both let us reduce subjectivity and make the impact of background noise/autofluorescence negligible.
Bleomycin is a radical-based mutagen that induces DSBs by the highly specific free radical attack on deoxyribose, in a very similar way to how low LET ionizing radiation (IR) acts. Thus, bleomycin is defined as a radiomimetic agent15. Since a great amount of bleomycin-induced DSBs could severely affect the proliferation of lymphocytes triggering, in the worst scenario, the activation of apoptosis, it is suggested to test several concentrations of the mutagen to identify a dose that can lead to DSBs formation without resulting in cytotoxicity.
Several studies show that the major pathway to repair IR-induced DSBs is NHEJ, which is promoted by the presence of 53BP1 near the lesion16,17. However, although a larger amount of 53BP1 foci might be expected to occur after treatment with bleomycin, which acts similarly to IR, in our study, we observed a higher frequency of γH2AX than 53BP1 foci. The main advantage of the γH2AX and 53BP1 foci analysis relies in the ability to evaluate the cellular response in either exposure or pathological contexts. In fact, it is well known that γH2AX and 53BP1 foci are reliable and established markers of IR effects18, as well as the presence of γH2AX foci represent an important prognostic factor in several forms of cancer19, or, more generally, they consent to determine the capability of agents or chemicals to increase DSBs occurrence. Furthermore, γH2AX and 53BP1 foci can be evaluated to establish a possible relationship between DSBs increase and a certain disease20,21, to verify a therapy in terms of response to the treatment22,23, or to estimate tumor radiosensitivity24.
γH2AX and 53BP1 foci assays are widely used in order to quantify DSBs, however, the authors must underline that both methods have limitations: the main concern consists in the fact that they do not detect DSBs themselves, but two specific factors involved in repair processes. Thus, considering that repair kinetics is highly variable25, the interpretation of the results could be ambiguous.
However, the present study indicates that the use of dual immunofluorescence to simultaneously quantify γH2AX and 53BP1 foci could lead to several issues in the interpretation of the results. This is mainly due to both the higher presence of γH2AX than 53BP1 at DSBs sites and the low levels of co-localized fluorescence signals for the two markers that we have observed after bleomycin treatment. On the other hand, histone H2AX remains phosphorylated on Ser-139 from the time a DSB appears until it is repaired, while 53BP1 is a protein that is recruited under more specific conditions, for example when only NHEJ can be activated, also, 53BP1 is strongly dominant during the G1 phase of cell cycle25. Furthermore, 53BP1 may have indeed been recruited where a DSB has occurred together with γH2AX, but persists for a shorter time, and therefore dual immunofluorescence is unable to localize it and γH2AX simultaneously. However, if the researcher deems it more appropriate to perform a dual immunofluorescence, for which we have provided a detailed and reliable protocol, it is important to consider that this method can inherently lead to underestimating the true frequency of the event, as described above. At the same time, in order not to overestimate the event, counting the co-localized γH2AX and 53BP1 foci as two different spots should be avoided since it indicates indeed the presence of only one DSB.
Divulgations
The authors have nothing to disclose.
Acknowledgements
We are grateful to the whole blood donors and all the health personnel who took the blood samples.
Materials
| AlexaFluor 568 goat anti-mouse IgG (γ1) | Invitrogen | A21124 | 53BP1 secondary antibody |
| Bleoprim | Sanofi | bleomycin sulfate (mutagen) | |
| Penicillin-streptomycin solution 100X | Euroclone | ECB3001D | antibiotics for culture medium |
| PBS 10X | Termofisher | 14200075 | Phosphate-buffered saline |
| FBS | Euroclone | EC20180L | Fetal Bovine Serum for immunofluorescence |
| Goat anti-rabbit IgG (H+L) DyLight 488 Coniugated | Termofisher | #35552 | γH2AX secondary antibody |
| Mouse anti-53BP1 monoclonal antibody | Merck | MAB 3802 | 53BP1 primary antibody |
| Labophot 2 | Nikon | Fluorescence microscope | |
| P-histone H2AX (Ser139) rabbit antibody | Cell Signaling | #2577 | γH2AX primary antibody |
| Phytohemoagglutinin | Termofisher | R30852801 | component of culture medium |
| Prolong gold antifade reagent with DAPI | Cell Signaling | #8961 | Antifade solution with DAPI for counterstaining |
| RPMI 1640 | Euroclone | ECB9006L | Culture medium |
| Triton-X100 | Sigma | T9284 | Nonionic detergent for permeabilization |
References
- Chatterjee, N., Walker, G. C. Mechanisms of DNA damage, repair and mutagenesis. Environmental and Molecular Mutagenesis. 58 (5), 235-263 (2017).
- Aleksandrov, R., Hristova, R., Stoynov, S., Gospodinov, A. The chromatin response to double-strand DNA breaks and their repair. Cells. 9 (8), 1853 (2020).
- Jackson, S. P., Bartek, J. The DNA-damage response in human biology and disease. Nature. 461 (7267), 1071-1078 (2009).
- Dickey, J. S., et al. H2AX: functional roles and potential applications. Chromosoma. 118 (6), 683-692 (2009).
- Her, J., Bunting, S. F. How cells ensure correct repair of DNA double-strand break. Journal of Biological Chemistry, Thematic Minireview. 293 (27), 10502-10511 (2018).
- Kuo, L. K., Yang, L. γ-H2AX – A novel biomarker for DNA double-strand breaks. In Vivo. 22 (3), 305-310 (2008).
- Bártová, E., Legartova, S., Dundr, M., Suchánková, J. A role of the 53BP1 protein in genome protection: structural and functional characteristics of 53BP1-dependent DNA repair. Aging. 11 (8), 2488-2511 (2019).
- Popp, H. D., Brendel, S., Hofman, W., Fabarius, A. Immunofluorescence microscopy of γH2AX and 53BP1 for analyzing the formation and repair of DNA double-strand breaks. Journal of Visualized Experiments. (129), 56617 (2017).
- Jezkova, L., et al. Particles with similar LET values generate DNA breaks of different complexity and reparability: a high-resolution microscopy analysis of γH2AX/53BP1 foci. Nanoscale. 10, 1162-1179 (2018).
- Scarpato, R., et al. Kinetics of nuclear phosphorylation (γ-H2AX) in human lymphocytes treated in vitro with UVB, bleomycin and mitomycin C. Mutagenesis. 28 (4), 465-473 (2013).
- Im, K., Mareninov, S., Diaz, M. F. P., Yong, W. H. An introduction to performing immunofluorescence staining. Methods in Molecular Biology. 1897, 299-311 (2019).
- Sanderson, M. J., Smith, I., Parker, I., Bootman, M. D. . Fluorescence microscopy. 10, (2014).
- Jamur, M. C., Oliver, C. Cell fixatives for immunostaining. Methods in Molecular Biology. , 55-61 (2010).
- Jamur, M. C., Oliver, C. Permeabilization of the cell membrane. Methods in Molecular Biology. 588, 63-66 (2010).
- Hecht, S. M. Bleomycin: New perspectives on the mechanism of action. Journal of Natural Products. 63, 158-168 (2000).
- Fei, P., El-Deiry, W. S. P53 and radiation responses. Oncogene. 22, 5774-5783 (2003).
- Mahaney, B. L., Meek, K., Lees-Miller, S. L. Repair of ionizing radiation-induced DNA double-strand breaks by non-homologous end-joining. Biochemical Journal. 417 (3), 639-650 (2009).
- Palla, V., et al. Gamma-H2AX: Can it be established as a classical cancer prognostic factor. Tumor Biology. 39 (3), 1010428317695931 (2017).
- Markovà, E., Hillert, L., Malmgren, L., Persson, B. R. R., Belyaev, I. Y. Microwaves from GSM mobile telephones affect 53BP1 and gamma-H2AX foci in human lymphocytes from hypersensitive and healthy persons. Environmental Health Perspectives. 113 (9), 1172-1177 (2005).
- Scarpato, R., et al. Nuclear damage in peripheral lymphocytes of obese and overweight Italian children as evaluated by the γ-H2AX focus assay and micronucleus test. The FASEB Journal. 25 (2), 685-693 (2018).
- Shanbhag, N. M., et al. Early neuronal accumulation of DNA double-strand breaks in Alzheimer’s disease. Acta Neuropathologica Communication. 7 (1), 77 (2019).
- Lassmann, M., et al. In vivo formation of gamma-H2AX and 53BP1 DNA repair foci in blood cells after radioiodine therapy of differentiated thyroid cancer. Journal of Nuclear Medicine. 51 (8), 1318-1325 (2010).
- Derlin, T., et al. Assessment of γ-H2AX and 53BP1 foci in peripheral blood lymphocytes to predict subclinical hematotoxicity and response in somatostatin receptor-targeted radionuclide therapy for advanced gastroenteropancreatic neuroendocrine tumors. Cancers (Basel). 13 (7), 1516 (2021).
- Djuzenova, C. S., et al. Radiosensitivity in breast cancer assessed by the histone γ-H2AX and 53BP1 foci. Radiation Oncology. 24, 8-98 (2013).
- Atkinson, J., Bezak, E., Kempson, I. Imaging DNA double-strand breaks – are we there yet. Nature Reviews in Molecular Cell Biology. 23, 579-580 (2022).
