Parts of this publication and its results have already been published (See references 3 and 23).
Experimental setup
An experimental setup is illustrated in Figure 1. A composite flow cycle comprises five flow chambers that are interconnected in series (Figure 1A). Within each flow chamber, four explants, each with a cross-sectional diameter of approximately 0.5 cm, are cultivated (Figure 1 A,C). For the static control experiment, the explants are cultivated in individual wells of a six-well plate (Figure 1B). To prevent the explants from being flushed out, they are affixed onto metal plates featuring narrow needle-shaped protrusions (Figure 1 D,E). In order to subject the explants to a direct flow of the medium, the chambers are inverted, with the inlets and outlets positioned at the head section (Figure 1 F,G). Within the bioreactor, the flow cycle is linked to a peristaltic pump. For the purpose of comparing tissue integrity between flow-cultured tissue and conventionally static-cultured tissue, explants are placed in a six-well plate adjacent to the flow cycle. This ensures the verification of consistent culture conditions in terms of oxygen, temperature, and humidity (Figure 1A)3.
Morphological analysis
β-actin
Various immunohistochemical staining procedures were conducted to examine histological distinctions in tissue integrity associated with diverse cultivation conditions (Figure 2). Explants that were promptly embedded post-dissection served as the baseline reference. For the analysis of the actin cytoskeleton within villous explants, β-actin staining was executed (Figure 2A–E). Descriptive analysis unveiled a well-structured and organized visual presentation of the cytoskeleton in freshly obtained tissue (Figure 2A). Over time, as cultivation progressed, there was an observable aggregation of microfilaments, signifying a degradation of the cytoskeletal structure. This phenomenon was consistently observed in villous explants that underwent static cultivation3 (Figure 2C,E, indicated by asterisks).
H&E staining
H&E staining provided additional reinforcement to the observation that tissue integrity diminishes over the course of static culture, a trend that is ameliorated in the context of flow culture (Figure 2F–J). Fresh tissue exhibited a structured and characteristic histological presentation of the villous explants, characterized by a dense and tightly packed stroma (Figure 2F). Additionally, the syncytiotrophoblast was firmly adhered to the underlying stroma (Figure 2F). A comparable appearance was noted in villous explants cultured in a flow environment for 24 h (Figure 2G). However, after 48 h of cultivation under the flow, portions of the syncytiotrophoblast were observed to be partially detached (Figure 2I, indicated by arrow), accompanied by sporadic small lacunae within the stroma. Histological scrutiny of the tissue indicated that the integrity of the tissue following 24 h in a static culture condition was inadequately preserved (Figure 2H). Furthermore, this integrity further degraded markedly after 48 h in static culture (Figure 2J). The stroma exhibited a porous and pitted appearance, and significant detachment of the syncytiotrophoblast from the stroma was evident in larger regions (Figure 2J, arrows)3.
CD34II
CD34II staining was employed to visualize endothelial cells and, consequently, the feto-placental blood vessels within the villous explants (Figure 2K–O). Tissue that was directly embedded right after dissection displayed a distinctively organized arrangement of the endothelial cells (Figure 2K). The morphological integrity of the feto-placental blood vessels remained well maintained after 24 h of flow culture and frequently even after 48 h, although occasional instances of collapsed blood vessels were noted under flow conditions (Figure 2 L,N). However, following 24 h of static culture, the blood vessels exhibited partial collapse, as evidenced by their disrupted visual appearance (Figure 2M, indicated by arrowheads). This deterioration of blood vessels within the static culture setting appeared to exacerbate with prolonged cultivation time. In summary, the descriptive morphological assessment of the villous explants subsequent to both flow and static culture indicated that tissue integrity appears to be more effectively preserved within the flow system when contrasted with the static culture mode3.
Ultrastructural analysis of the cultivated tissue
Transmission electron microscopy
To conduct a more detailed examination of the morphology of the villous explants, additional ultrastructural analyses were performed using transmission electron microscopy (TEM) (Figure 3A–E). These findings corroborated the outcomes of the histological investigations. In tissue that was directly embedded immediately following preparation, the morphology was exceptionally well preserved (Figure 3A). Microvilli were clearly discernible on the surface of the syncytiotrophoblast. The syncytiotrophoblast presented its distinctive continuous layer without lateral cell boundaries, establishing direct contact with the basement membrane. The stroma of the fresh tissue exhibited dense packing without significant perforations or ruptures. Moreover, the ultrastructural appearance of the blood vessels and individualized intravascular erythrocytes also demonstrated excellent preservation (Figure 3A).
Even after 24 h of flow culture, the overall morphology of the tissue samples remained relatively well maintained (Figure 3D). While there were slightly fewer microvilli on the surface of the syncytiotrophoblast compared to fresh tissue, the syncytiotrophoblast remained primarily attached to the basal membrane. Nuclei and occasional small vacuoles were observable within the inner portion of the syncytiotrophoblast. The stroma within the placental villi appeared well-preserved and closely resembled fresh tissue (Figure 3D). Even following 48 h of flow culture, the stromal cells exhibited relatively good preservation, albeit with some perforations present (Figure 3E). Intriguingly, lipid droplets were detected within the tissue. While the syncytiotrophoblast displayed vacuoles and a reduction in the number of microvilli, it remained attached to the basal membrane in numerous regions, and syncytial and cell nuclei were clearly visible (Figure 3E).
In stark contrast to the tissue from the flow culture, the morphology of villous tissue subjected to static culture exhibited deterioration as early as 24 h (Figure 3B). The syncytiotrophoblast became dissociated from the basal membrane at multiple sites and displayed relatively substantial perforations. Additionally, lipid droplets were frequently evident in both the syncytiotrophoblast and the stroma (Figure 3B). Following 48 h of static culture, a progressive decline in ultrastructure was apparent (Figure 3C). The syncytiotrophoblast presented numerous perforations and detachment from the basal membrane to a considerable extent. Identifying cells within the stroma, as well as endothelial cells comprising the blood vessels, became challenging. Furthermore, there was a notable accumulation of lipid droplets within the villous explants after 48 h of static culture (Figure 3C). In summary, the ultrastructure of tissue in static culture exhibited successive deterioration over the duration of the cultivation period, a trend that was mitigated by cultivation under flow conditions3.
Scanning electron microscopy
Utilizing scanning electron microscopy (SEM), a detailed examination of the surface of the villous explants was facilitated (Figure 4A–J). Tissue that had been freshly embedded exhibited a densely populated array of microvilli across its surface (Figure 4 A,B). Some regions exhibited vesicle-like structures. In contrast, tissue from the static culture manifested a substantial reduction in microvilli after 24 h (Figure 4C,D), a reduction that persisted after 48 h (Figure 4E,F). While certain areas showed aggregation of vesicle-like structures that had not been released, other regions appeared bare and eroded (Figure 4D,F). In tissue subjected to flow culture, microvilli were still present on the surface after 24 h (Figure 4G,H), as well as after 48 h (Figure 4I,J), although to a lesser extent than in fresh tissue. In comparison to the static culture, the prevalence of vesicle-like structures on the surface was diminished. Intriguingly, these vesicle-like structures were notably concentrated in specific recesses where flow could be reduced or absent (Figure 4H,J), suggesting that they might have been dislodged from the flow-exposed tissue surface due to the flow of the medium3.

Figure 1: Set up of the flow system. (A) The assembled flow system, consisting of the reservoir and five flow chambers, is connected to one of the peristaltic pumps. On the right side is a six-well plate in which the explants are statically cultured. (B,C) For both cultivation methods, the placental samples are dissected into villous explants of approximately 0.5 cm2, of which four explants are then used per well or chamber. In an experimental approach, five chambers or wells are used. (D,E) For flow culture, a metal plate with narrow needle-shaped elevations is used to secure the explants. (F,G) The openings of the tubes are located at the head of the chambers, and are thus used upside down to guarantee that the tissue is exposed to direct flow. This figure is reproduced from Kupper et al.3. Please click here to view a larger version of this figure.
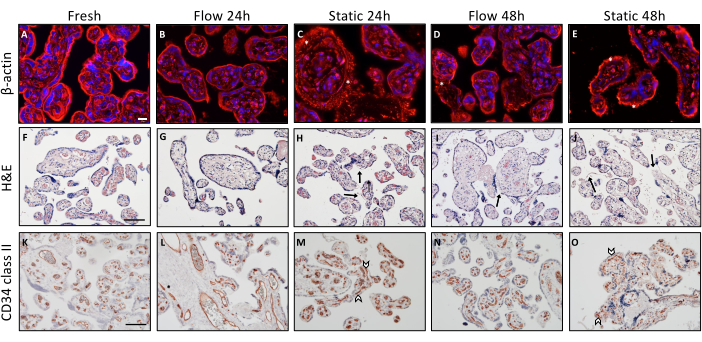
Figure 2: Morphological analysis of placental villous explants upon flow and static culture. (A–E) Immunofluorescence staining for β-actin to visualize the cytoskeleton of explants upon culture. For the analysis, six randomly selected spots were used per slide. Representative images are shown. (A) Visualization of the cytoskeleton of tissue embedded directly after preparation. Scale bar: 20 µm. (B–E) Representative depiction of both time-dependent and cultivation mode-dependent degeneration of the actin cytoskeleton in cultured explants of flow and static culture. (C–E) Asterisks signify increased actin microfilament accumulation, which is an indication of actin cytoskeleton degradation. (F–J) Hematoxylin-eosin staining of villous explants. Scale bar: 100 µm. (F,G) Freshly embedded tissue (F) and flow culture explants for 24 h (G) show a well-preserved morphology of a villous explant. (I) Flow-cultured explants for 48 h show intermittently detached areas of the syncytiotrophoblast (arrow). (H,J) Time-dependent deterioration of structural integrity after static explant culture, indicated by dislodgment of the syncytiotrophoblast (arrow) and perforated stroma. (K–O) CD34 II was used to stain villous endothelial cells. Scale bar: 100 µm. (K,L) Fresh tissue (K) and explants cultured for 24 h under flow conditions (L) exhibit a characteristic structurally aligned endothelial cell pattern. (N) After 48 h in flow culture, the vascular integrity decreases to some extent. (M,O) In static culture, collapsed blood vessels are already visible after 24 h (M), which was observed to increase with longer static cultivation time (O). This figure is reproduced from Kupper et al.3. Please click here to view a larger version of this figure.
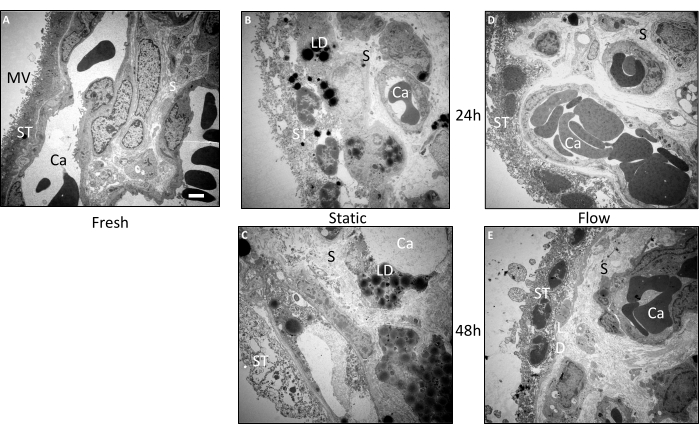
Figure 3: Ultrastructural pre- and post-cultivation examination of villous explants using transmission electron microscopy. Tissue from three independent experiments was used to analyze the images. (A) A representative image of freshly embedded tissue shows a large amount of microvilli (MV) on the surface of the syncytiotrophoblast (ST). Structurally intact capillaries (Ca) are visible in the well-preserved stroma (S). (B) In the tissue that has been statically cultured for 24 h, there is a deterioration in the structural integrity of the syncytiotrophoblast, which appears to be disconnected from the basal membrane in some areas. There is also a noticeable accumulation of lipid droplets (LD). (C) After 48 h in static culture, severe ultrastructural deterioration is observed. The stroma as well as the syncytiotrophoblast are perforated and a massive accumulation of lipid droplets is evident. Blood vessels could scarcely be traced. (D,E) The ultrastructure of the tissue from the flow culture was relatively well preserved after 24 h (D) as well as after 48 h (E). Scale bar: 2 µm. MV: Microvilli, ST: Syncytiotrophoblast, S: Stroma, Ca: Capillary, LD: Lipid droplets. This figure is reproduced from Kupper et al.3. Please click here to view a larger version of this figure.

Figure 4: Ultrastructural pre- and post-cultivation examination of villous explants using scanning electron microscopy. (A,C,E,G,I) Overview images of the surface of the placental villous trees with respective detailed images (B,D,F,H,J). (A,B) Freshly embedded tissue exhibits a dense seam of microvilli. (B) Vesicular-like structures can be noticed in some locations. (C–F) After 24 h and 48 h in static culture, a decrease in microvilli on the surface of the syncytiotrophoblast is visible. Striking is the extensive accumulation of vesicular-like particles on the surface of the explant. (F) The particles appear to wither after 48 h in static culture. (G–J) The surface of the tissue from the flow culture seems to be better preserved after 24 h (G,H) as well as after 48 h (I,J) compared to the static culture. Microvilli are visible on the surface (H,J), although not in the same high density as in the fresh tissue. (B) Vesicular particles can be seen scattered in the niches with reduced flow. This figure is reproduced from Kupper et al.3. Please click here to view a larger version of this figure.
Supplementary Table 1: Experimental settings for placental villous flow and static culture. Please click here to download this File.
Supplementary Table 2: Specifications of the flow and static system. Please click here to download this File.
Supplementary Table 3: Antibodies for immunohistochemistry and immunofluorescence used for this study. Please click here to download this File.
