These protocols yield reproducible stromal cell varieties from the thymus and bone marrow suitable for flow cytometric analysis, as well as single-cell multiomics, such as scRNA sequencing. Murine thymic tissue undergoes significant remodeling in response to stressors, such as the cytotoxic conditioning that precedes bone marrow transplantation or the natural aging process. As a consequence, thymic cellularity is drastically reduced in both of these settings (Figure 1A). While a thymus from an 8-week-old wild-type mouse contains approximately 100 million cells, the cellularity of a 2-year-old mouse can be expected to be half of that, and a thymus 4 days post-irradiation and bone marrow transplantation can be as low as 5,00,000 cells (Figure 1A).
Cytotoxic conditioning will preferentially ablate hematopoietic cells, so the stromal compartment will be proportionally enriched (Figure 1B). However, the overall low cellularity will make isolating sufficient numbers of thymic stromal cells challenging. It may, therefore, be necessary to pool animals to obtain cell numbers compatible with downstream multiomics applications and to minimize the loss of material wherever possible. To perform multiomics on thymic stromal cells, fluorescence-activated cell sorting (FACS) of live, single, CD45-Ter119- cells (Figure 1C) will ensure that sufficient numbers of cells can be sorted from most stress conditions.
To preserve the heterogeneity of the stroma compartment, we typically do not sort on a single positive stroma marker but gate out hematopoietic cells. Including positive stromal cell markers is still recommended for analysis to ensure that proper tissue dissociation has been achieved. However, FACS isolation is imperfect, and as determined by scRNAseq, 30%-50% of the sorted cells will be of hematopoietic origin (Figure 1D,E). This can be improved upon by magnetically depleting CD45+ cells prior to FACS; however, this is not recommended for conditions of reduced thymic cellularity (Figure 1A) as the loss of material that comes with the extra processing involved in magnetic depletion results in stroma yields too low for downstream applications.
For the isolation of human thymic stroma, the most critical step is the enzymatic digestion. If the dissociation step is rushed, the overall stromal cell yield, as well as stromal cell subtype representation, will be severely diminished (Figure 2A). It is also recommended to always include the markers of thymic endothelium and epithelium to ensure that tissue digestion has been optimal, as simply gating negative for hematopoietic markers will not be sufficient to gauge actual stromal cell release. This is exemplified in Figure 2A, where the percentage of cells negative for hematopoietic markers is higher in the sample digested for 30 min compared to the sample incubated for 90 min. However, as seen in the consecutive gate, the yield of endothelial and epithelial cells is very poor in the shorter digestion protocol (Figure 2A).
For FACS isolation of human thymic stromal cells for multiomics applications, staining with a hematopoietic lineage antibody cocktail, as well as CD4 and CD8 antibodies is suggested, in addition to the pan-hematopoietic marker CD45 and the erythroid marker CD235a (Figure 2B) to improve stromal cell enrichment. Thus, for human multiomics analysis of thymic stroma, live, single, CD45-CD235a-Lineage-CD4-CD8- cells should be FACS sorted (Figure 2B). As seen for scRNAseq of murine samples, this still results in contamination of 30%-50% hematopoietic cells, but this still gives acceptable resolution of the human thymus stromal compartment (Figure 2C,D).
For bone and BM stroma, the same strategy was used as for thymic tissue; stromal cells are isolated by gating on cells negative for hematopoietic markers, and further stromal cell markers are included as digestion controls (Figure 3). The dissociation of calcified tissue like bone results in a significant accumulation of debris. The addition of calcein as a viability dye will allow the exclusion of most of this debris, which otherwise falsely inflates sorted numbers and decreases gated population frequencies (Figure 3). This is exemplified in the FACS plots comparing gating on all cells and just gating on Calcein+ (Figure 3). As seen for human thymic stromal cells, the percentage of hematopoietic marker-negative cells is not a reliable readout for bone and BM tissue dissociation efficiency (Figure 3). While the CD45-Ter119- gate can contain an abundance of cells in a sample where the digestion step failed, further analysis of typical bone and BM stromal cell markers CD31, CD140a, and CD51 clearly demonstrate poor stroma release (Figure 3 and Figure 4).
For FACS isolation of murine bone and BM stromal cells, using the gating strategy is suggested as in Figure 3, where we only sort CD45-Ter119-Calcein+ but monitor the forward and side scatter properties and stromal cell marker expression of the sorted population to ensure optimal digestion (Figure 3A). The combination of hematopoietic lineage-positive magnetic cell depletion of the bone marrow prior to FACS and sorting on the shown gates results in single-cell sequencing of cells that are >90% stroma. An example of such a scRNA seq run (Figure 5A,B) illustrates this low hematopoietic contamination after transcriptome-based annotation of the analyzed cells (examples of hematopoietic marker genes Figure 5C). This is in stark contrast to the thymic stromal preparations where magnetic cell depletion was not performed before the sorting step due to the limited starting material in post-transplant thymic tissue (Figure 1F).
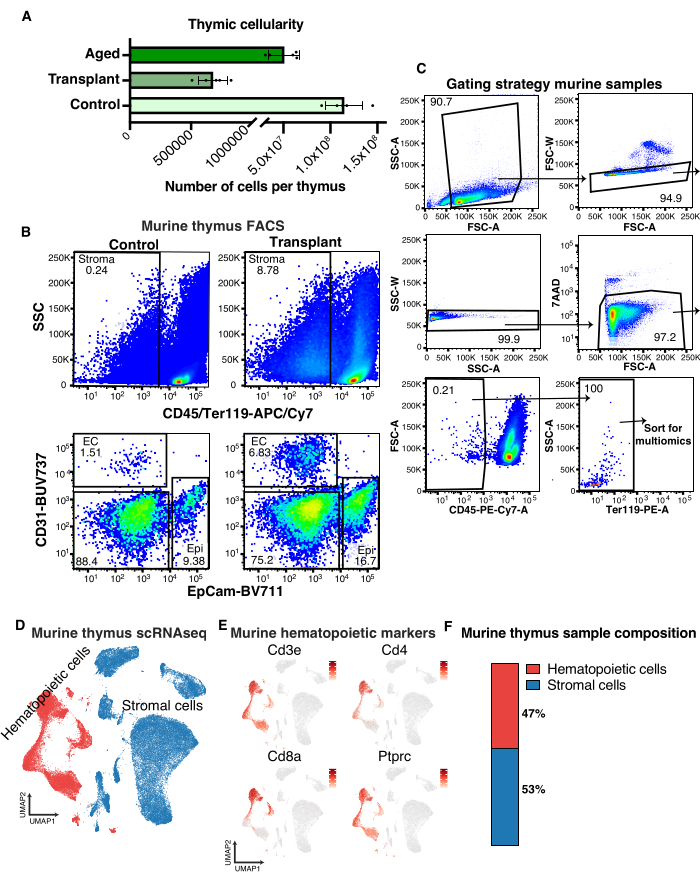
Figure 1: Flow cytometric and single-cell RNA sequencing analysis of murine thymic stroma. (A) Quantification of thymocyte numbers in 8-week-old mice (Control), 8-week-old mice lethally irradiated and transplanted 4 days prior (Transplant) and 2-year-old mice (Aged). (B) Representative flow cytometry plots demonstrating the enrichment in stromal cell subtypes 4 days following lethal irradiation and transplantation. (C) Flow cytometry gating strategy to sort murine thymic stroma for single-cell multiomics analysis. (D) UMAP of murine thymus indicating stroma (blue) and hematopoietic cells (red). (E) UMAPs showing murine hematopoietic marker genes in thymus samples. (F) Quantification of murine stromal and hematopoietic cells in thymus scRNAseq samples. Please click here to view a larger version of this figure.
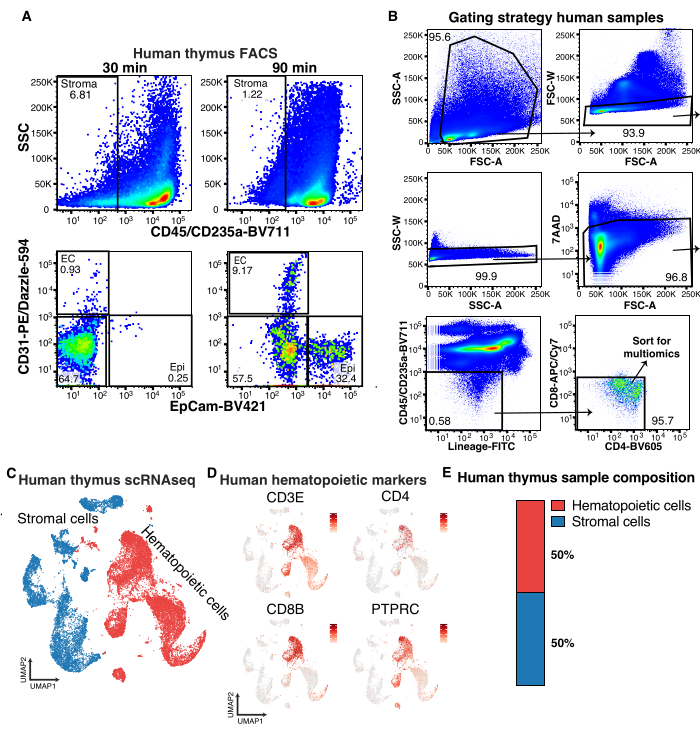
Figure 2: Flow cytometric and single-cell RNA sequencing analysis of human thymic stroma. (A) FACS plots showing representative results of the impact of different digestion times on the release of human thymic stromal cell populations. (B) Flow cytometry gating strategy to sort human thymic stroma for single-cell multiomics analysis. (C) UMAP of human thymus indicating stroma (blue) and hematopoietic cells (red). (D) UMAPs showing human hematopoietic marker genes in thymus samples. (E) Quantification of human stromal and hematopoietic cells in thymus scRNAseq samples. Please click here to view a larger version of this figure.
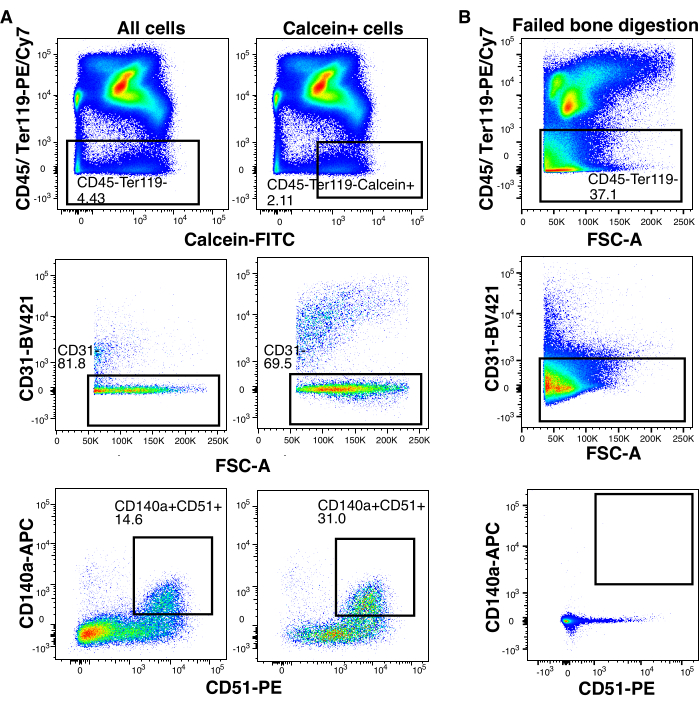
Figure 3: Murine bone marrow stromal markers and Calcein are helpful to estimate dissociation efficiency. (A) Representative flow cytometry analysis of CD45–Ter119– murine bone marrow stromal cells gated on all cells and Calcein+ cells. With subsequent CD31– gating showing a higher proportion of CD140a+CD51+ cells when selecting Calcein+ cells. (B) Flow cytometry analysis representative of a failed bone digestion with missing CD140a+CD51+ cells. Please click here to view a larger version of this figure.
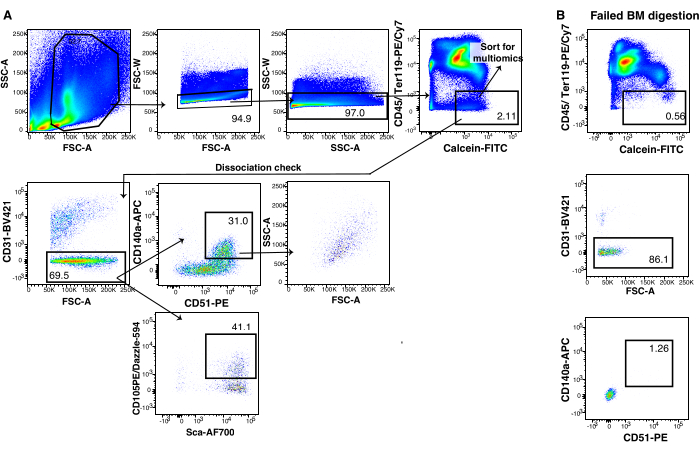
Figure 4: Bone marrow stroma gating strategy. (A) Flow cytometry gating strategy to sort murine bone marrow stroma for single-cell multiomics. (B) Flow cytometry analysis representative of a failed bone marrow digestion with missing CD140a+CD51+ cells. Please click here to view a larger version of this figure.
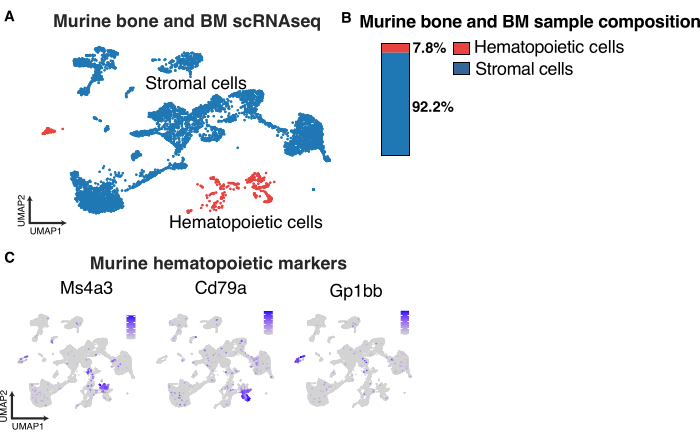
Figure 5: Bone marrow single-cell RNA sequencing with stromal and hematopoietic cell composition. (A) UMAP of murine bone and bone marrow indicating stroma (blue) and hematopoietic cells (red). (B) Quantification of stromal and hematopoietic cells in murine bone and bone marrow samples. (C) UMAP of hematopoietic marker genes in murine bone marrow samples. Please click here to view a larger version of this figure.
