Cell aggregation is important for promoting cell-to-cell contact and signaling. Encasing cell aggregates inside the basement membrane matrix supported both 3-dimensional cultures for in vitro tissue organoid formation and facilitated the mechanical delivery of cells into the kidney capsule for graft transplantation. To establish these constructs, the basement membrane matrix was first maintained in a fluidic state under ice-cold conditions. Cell aggregation was subsequently achieved by layering a concentrated cell suspension above and using centrifugal force to push cells through the high-density matrix. Optimization of the centrifugation speed (200-2000 x g) was required to achieve a mid-layer cell positioning (Figure 1A), which was also dependent on the cell number. Higher centrifugal forces (>500 x g) propelled cells to the very tip of the pipette (Figure 1B), and caution is required for smaller cell numbers (i.e., <5 x 105) that may be lost during the removal of the flexible laboratory film covering. Conversely, cells may not travel adequately through the basement membrane matrix if G-forces are too low (≤200 x g) (Figure 1C). Centrifugation speed for cell numbers <1 x 105 or >2.5 x 106 may need further optimization. Following centrifugation, cells were set inside the basement membrane matrix by warming the construct at 37 °C for 15 min. This solidification process enabled the matrix "plug" to be ejected entirely (Figure 1D) using a thin wire plunger.
In vitro spleen organoid formation
Basement membrane matrix plugs ejected into an appropriate tissue culture vessel reproducibly formed 3-dimensional organoid-like structures which could be maintained following standard tissue culture techniques and conditions (37 °C, 5% CO2, 95% humidity)11. In culture, the supporting basement membrane matrix substrate gradually dissipated between days 0 and 14 (Figure 2A, i-iii) and could no longer be observed by day 21, leaving an intact organoid-like spherical cell mass (Figure 2A, vi) measuring approximately 545 µm (s.d. = 120 µm, n = 6)11 in diameter. These organoid structures were supported in culture beyond 30 days but did not increase in size over time (Figure 2B). Spleen organoids were composed of CD45–TER-119– stromal, CD45–TER-119+ erythrocyte, and CD45+TER-119– lymphoid cells (Figure 2C), however, the frequency of each population was variable across individual organoids (n = 4; Table 1). To assess the general structure of spleen organoids, 50 µm thick tissue cryosections were prepared and visualized. Organoids were comprised of a non-hollow structure, with cells present across the entire diameter of the tissue (Figure 2D). Assessment of organoid sections at 7 µm single cell layer thickness revealed that cells were arranged in cord-like structures without a clear spatial orientation (Figure 2D), with areas void of nuclei between strings of cells. CD45 antibody staining verified the presence of nucleated hematopoietic cells which densely surrounded peripheral regions of the organoid (Figure 2E). In addition, CD45+ cells of distinct, spindle-shaped morphology were observed in more central organoid regions. A general absence of CD90.2 (T cell) and CD19 (B cell) antibody staining, but positive CD11b staining, demonstrated the specific presence of myeloid cells (Figure 2F). Non-hematopoietic CD105+ and CD31+ endothelial cells were also detected in multiple organoids11.
In vivo spleen graft transplantation
The encapsulation of aggregated spleen stromal cells within a semi-solid matrix facilitates animal transplantation studies. This technique was used to embed unfractionated or CD45–TER-119– FACS sorted neonatal spleen stromal cell preparations inside a basement membrane matrix plug, serving as a vehicle for transplantation under the mouse kidney capsule. In line with similar cell-aggregation protocols8, basement membrane matrix encapsulated cell constructs (MECCs) successfully regenerated spleen tissue in 4/5 independent animal transplantations, thereby confirming the viability of this graft construction technique. To test the utility of MECCs in defining cell types required for spleen regeneration, MECC grafts were constructed from neonatal spleen cells which were specifically depleted of PDGFRβ+MAdCAM-1+ or PDGFRβ+MAdCAM-1– stromal (CD45–) cells11. Grafts lacking PDGFRβ+MAdCAM-1+ cells retained the capacity for gross tissue regeneration (4/4 grafts; Figure 3A)11. Three out of four regenerated tissues exhibited normal spleen cell composition and structure, displaying central arterioles, white pulp follicles, segregated T and B cell compartments, follicular dendritic cells, marginal zone reticular cells, and marginal metallophilic macrophages, red pulp sinusoids, myeloid cells and macrophages (Figure 3B)11. In contrast, grafts that lacked PDGFRβ+MAdCAM-1– cells largely failed to regenerate spleen tissue (1/4 grafts)11. These data support the importance of graft-derived PDGFRβ+ cells in spleen tissue regeneration8, and pinpoint a specific requirement for PDGFRβ+MAdCAM-1– stromal cells.
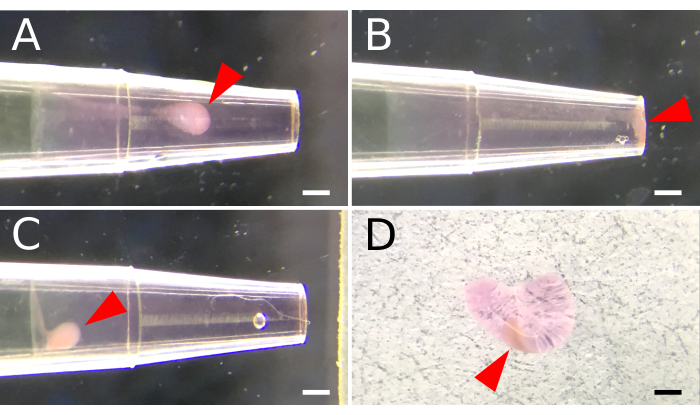
Figure 1. Basement membrane matrix encapsulation of aggregated spleen stromal cells. A 200 µL pipette tip is used to facilitate cell aggregation inside a basement membrane matrix. A fluidic matrix layer is first established by aspirating 2 µL of ice-cold basement membrane matrix into a pre-chilled pipette tip. A cell suspension is deposited above the matrix layer, and centrifugal force is applied to mobilize and aggregate cells within the basement membrane matrix. (A) Settings for centrifuge speed are optimized to position cell aggregates midway along the matrix layer. (B) Excessive centrifugal speed results in cells that aggregate at the end of the pipette. (C) Insufficient speed prohibits cell movement through the matrix. (D) A semi-solid matrix plug is ejected from the pipette tip following incubation at 37 °C for 15 minutes. Arrowheads indicate the placement of cell aggregates within the matrix. Scale Bar, 100 µm. Please click here to view a larger version of this figure.
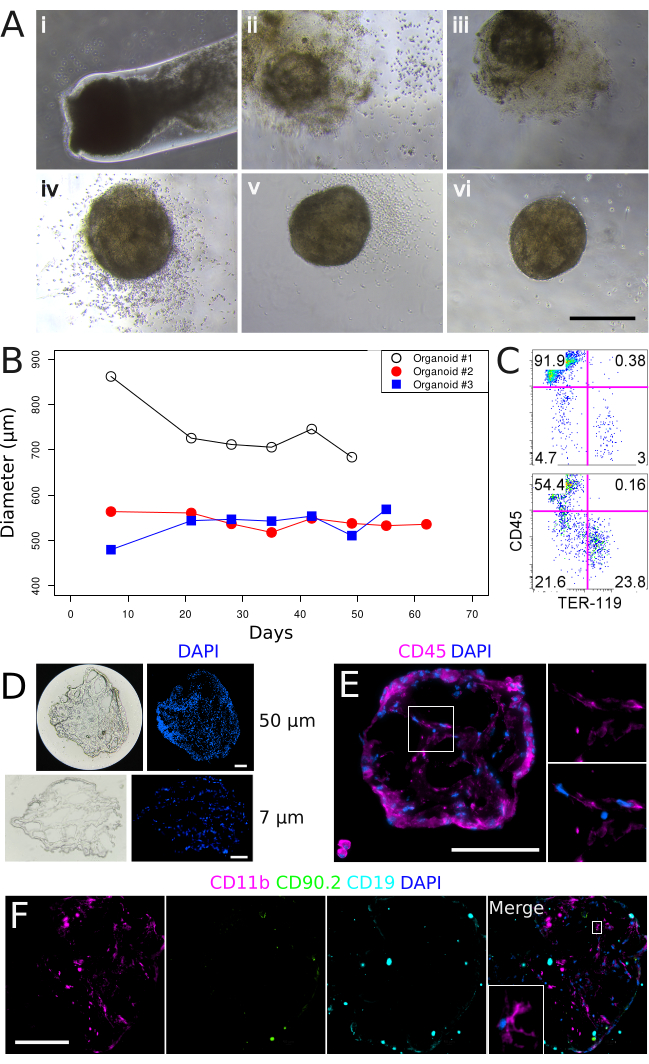
Figure 2. In vitro culture of basement membrane matrix-encapsulated spleen aggregates and the formation of 3-dimensional organoid structures. (A) Spleen organoid development over a 35-day time course. Panels (i-vi) correspond to Days 0, 7, 14, 21, 28, and 35 in culture. Images were captured on a Nikon TS2 Inverted Phase Contrast Microscope. Scale bar, 500 µm. (B) Spleen organoid diameter over extended culture periods. Each line represents an individual organoid. (C) Flow cytometry characterization of stromal (CD45–TER-119–), lymphoid (CD45+TER-119–) and erythroid (CD45–TER-119+) cell populations after 53 days in culture. Upper panel: Control tissue prepared from neonatal spleen stroma. Lower panel: Spleen organoid, Day 52. (D) Organoid tissue gross morphology at 50 µm (upper panels) and 7 µm (lower panels) section thicknesses after 22 days in culture. (E) Localization and morphology of CD45+ hematopoietic cells after 34 days in culture. Magnified region is shown in right panels. Sections are 7 µm. (F) Assessment of myeloid (CD11b), T (CD90.2) and B (CD19) cells after 34 days in culture. Magnified region is shown in inset. Sections are 7 µm. Images were captured using a Nikon Eclipse Ti2-E Live Cell Microscope. Scale bars (D, E, F), 100 µm. Please click here to view a larger version of this figure.
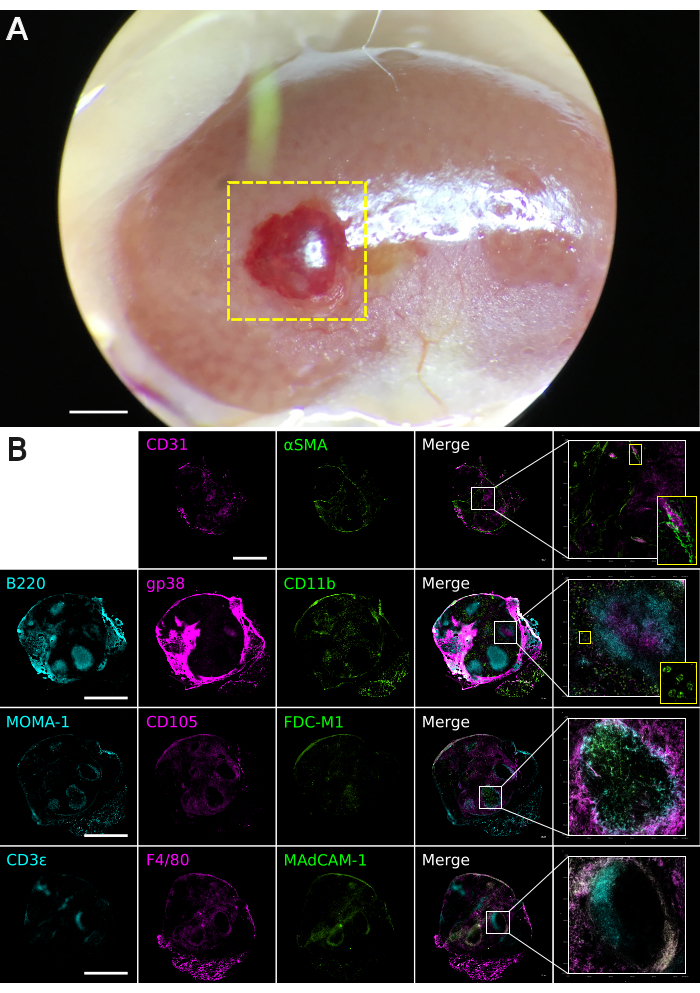
Figure 3. Basement membrane matrix graft tissue analysis. Grafts were constructed from neonatal spleen stroma depleted of PDGFRβ+MAdCAM-1+ cells (n = 4). Additional data available online11. (A) Macroscopic appearance of a regenerated spleen graft at 4 weeks post-transplantation under the kidney capsule. The boxed yellow area indicates regenerated spleen tissue. The image was captured using a Leica M60 Stereomicroscope equipped with a Snap zoom adapter and an Apple iPhone 6S. Scale bar, 1000 µm. (B) Composite multi-color immunofluorescence images of 30 µm-thickness graft tissues cryosectioned on the coronal plane. Antibody staining was performed with indicated markers to visualize spleen tissue micro-architecture. Images were captured using a Nikon Eclipse Ti2-E Live Cell Microscope. Boxed areas show regions of higher magnification. Scale bar, 1000 µm. Please click here to view a larger version of this figure.
| Organoid No. | Stromal cells | Erythrocytes | Lymphocytes |
| 1 | 62 | 7.7 | 31 |
| 2 | 22 | 24 | 55 |
| 3 | 3.6 | 0.1 | 96 |
| 4 | 10 | 76 | 12 |
Table 1. Percentage of stromal, erythroid and lymphoid cell populations amongst individual spleen organoids.
Supplementary figure 1. Schematic overview of the protocol highlighting the major steps in generating matrix embedded cell constructs for in vitro and in vivo studies. Please click here to download this File.
