The sections for the molecular machine-based detection should be prepared first because their preparation takes more time than the assembly of the molecular device. The construct works well with 5-6μm-thick sections cut from paraformaldehyde-fixed, paraffin-embedded tissue blocks. Use slide brands which retain sections well, such as ProbeOn Plus charged and precleaned slides (Fisher Scientific) or similar. We recommend at first using a tissue with a well-known pattern of DNA damage which contains both DNase I- and DNase II-type breaks, such as dexamethasone-treated apoptotic rat thymus 13,14.
1. Preparation of sections
- Place the sections in a slide rack and dewax in xylene for 15 min, transfer to a fresh xylene bath for an additional 5 minutes.
- Rehydrate by passing through graded ethanol concentrations: 96% Ethanol -2x5min; 80% Ethanol – 5min; water – 2×5 min.
- Digest section with Proteinase K. Use 100μL of a 50μg/mL solution per section. Incubate 15′ at room temperature (23° C) in a humidified chamber. The time may need adjustment depending on the tissue type. Hard tissues might require longer digestion. Times of 15-25 min are usually used. Insufficient digestion may result in the weaker signal. Overdigestion on the other hand results in signal disappearance and section disruption.
- Rinse in distilled water for 2×10 min.
- Apply 100 μL per section of 2% BSA for preblocking. Incubate for 15 min at room temperature (23° C). During this time assemble the molecular machine.
2. Molecular machines assembly
All reagents are scaled for 25 μL total volume, which is sufficient for a single detection in an average size tissue section (10x10mm). The volume can be scaled up as needed.
- Combine in a small plastic tube in this order:
|
- Mix gently by pipetting. The molecular construct almost instantly self-assembles in this solution and can be used immediately at room temperature (23° C). Increasing the temperature to 37° C blocks the T4 DNA ligase-based component of the labeling.
3. Using molecular machines in tissue sections to dual label 5’OH and 5’PO4 DNA breaks
- Aspirate the preblocking solution and apply 25μL of the full reaction mix containing 70 pmoles of Oligonucleotide 1, 53 – 215 pmoles (1.76 – 7.1 μg) VACC TOPO and 10 units T4 DNA ligase (500 U/mL) in solution of 66 mM-Tris HCl, pH 7.5, 5mM MgCl2, 0.1 mM dithioerythritol, 1 mM ATP, and 15% polyethylene glycol-8000. The same sequence oligonucleotide carrying a single FITC label, Oligonucleotide 2, can be used instead for detection of a single type of DNA breaks. In such case omit ligase from the labeling reaction. Also in this situation a solution of 50mM Tris-HCL, pH 7.4, 15% PEG-8000 can be used instead of T4 DNA ligase buffer.
- Incubate for 1 hr at room temperature (23° C) in a humidified chamber with a plastic coverslip. Protect from light. Lowering of the temperature to 16° C reduces the ligase-based signal; the temperature increase to 37° C completely eliminates the ligase-based signal. A partial inhibition of ligase sometimes occurs in the reaction mix, possibly due to contaminants introduced with topoisomerase preparations. Therefore, longer incubation (2-4 hours) might be required especially when significant numbers of ligase-labeled breaks are present. Although both ligase and topo signals can be observed at this stage, in many instances the ligase signal can be further enhanced by re-application of the reaction mix without VACC TOPO and dual-labeled Oligonucleotide, but containing a hairpin shaped oligonucleotide (Oligonucleotide 3) (35 mg/mL) and T4 DNA ligase (250 U/mL). To enhance signal proceed to Step 3, to see the reaction without enhancement go to Step 5.
- Remove coverslips by gently immersing the slides vertically in a Coplin jar containing water at room temperature. Aspirate excess water.
- Enhance ligase signal by applying 25 μL of reaction mix without VACC TOPO and Oligonucleotide 1, but containing Oligonucleotide 3: 66 mM-Tris HCl, pH 7.5, 5 mM MgCl2, 0.1 mM dithioerythritol, 1 mM ATP, and 15% polyethylene glycol-8000, 5 units T4 DNA ligase, 35 mg/mL Oligonucleotide 3 (blunt ended hairpin). The total volume of the labeling solution can be scaled up to accommodate the bigger sections. Incubate for 18 hr (overnight) at room temperature (23° C) in a humidified chamber with a plastic coverslip.
- Remove coverslips by gently immersing the slides vertically in Coplin jar containing water at room temperature. Then wash section 3×10 min in distilled water.
- Rinse with sodium bicarbonate buffer. Alkaline solution rinse enhances FITC fluorescence, which is pH sensitive and is significantly reduced below pH 7.
- Cover section with an antifading solution (Vectashield with DAPI), coverslip and analyze the signal using a fluorescent microscope. Double-strand DNA breaks with 5’OH will fluoresce green, 5’PO4 breaks – will fluoresce red (Figure 2).
4. Representative Results
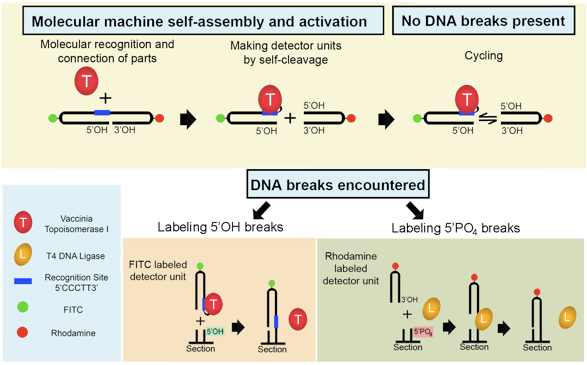
Figure 1. Semi-artificial molecular machine detects two types of DNA damage in situ. The fluorescent machine self-assembles when VACC TOPO binds to the double-hairpin 32-mer. The machine begins operation by splitting itself into two detector units via topoisomerase-made cut at the 3′ end of the recognition sequence. This results in a cyclic process where the FITC-labeled unit continuously separates and religates back to the rhodamine-labeled unit. This persists until a detectable DNA break is encountered. When such an alternative acceptor (5’OH blunt end DNA break) is present in the tissue section, the FITC part will ligate to it. The remaining rhodamine part will attach to 5′ PO4 DNA blunt end breaks with the help of T4 DNA ligase. Consequently, both types of DNA breaks are simultaneously detected.
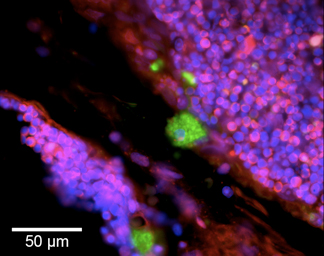
Figure 2. Molecular machine dual labels two types of DNA breaks in a tissue section of dexamethasone-treated thymus. Blunt-ended DNA breaks of DNase I- and DNase II-type are detected in the thymic cortical areas undergoing apoptosis. Green cytoplasmic fluorescence (5’OH DNA breaks) marks cortical macrophages digesting nuclear material of apoptotic thymocytes. This signal is produced by VACC TOPO, and localizes to phagolysosomes with DNA containing 5’OH double-strand breaks 11,12. Red fluorescence (5’PO4 breaks) labels nuclei of apoptotic thymocytes not engulfed by macrophages. Massive numbers of thymocytes simultaneously undergo apoptosis accompanied by generation of 5’PO4 double-strand breaks, visualized by ligase-based labeling. These breaks are located at the nuclear periphery, forming ring-shaped patterns. All cellular nuclei are visualized by counterstaining with DAPI (blue fluorescence).
