1. Sample Acquisition
- Choose and prepare the sampling site one day prior to sampling the water column. This will allow the stratified layers of the water column to reform after disturbance due to drilling and ice-hole melting. Identify location of drill site by GPS.
- To access the water column, begin by drilling a hole through the ice with a Jiffy ice auger attached to a 4-inch Jiffy flight extension and cutting bit. To prevent freezing of the drill in the hole, try to avoid drilling into the liquid water by halting drilling approximately 4-6 inches above the top of the water column.
- The drill hole will need to be widened from a diameter of 15 to 60 cm prior to sampling. This is accomplished by use of a hole melter (Hotsy Model) which circulates heated polyethylene glycol through a copper pipe. Hole melting typically takes up to 18 hours.
- Prior to sampling the water column, ensure that all required equipment and sample storage vessels are assembled at the sampling site. Sampling materials include: Niskin bottle (5-10 L capacity), Winch with depth-calibrated cable, Messenger, sufficient sample bottles for each sampling depth and coolers for sample transport.
- Begin sampling early in the day (~5 am), as lake water filtering should be completed on the day of sampling. Collect water samples (1-5 L) from desired depths (for this study: 6-m, 15-m and 18-m sampling depths, measured from the piezometric water level in the ice hole) using a Niskin sampler attached to a calibrated hand winch. To prevent disturbance of the water column collect shallow depths before deeper ones.
- Store lake samples in coolers and transport from the sample site to the field lab using a sled attached to an ATV (all terrain vehicle). Samples should be processed or stabilized (for eg: flash freeze in liquid nitrogen) at the field laboratory immediately following sampling.
2. Development of Enrichment Cultures
- For cultivation of enrichment cultures,Concentrate 0.5-1 L of lake water onto 47 mm 0.45 μm pore size polyethersulfone filters using gentle filtration (< 0.3 atm). If the diversity of the natural community is desired, filter a second sample onto 47 mm 0.45 μm pore size polyethersulfone filters and follow protocols according to Bielewicz et al. 4 for eukaryote phylogenetic diversity.
- Transfer filter to 50 mL sterile falcon tube containing ~20 mL of filtered lake water collected from the same sampling depth as the filter. Gently wash cells from the filter using a sterile pipette.
- Transfer cell suspension to a 25 cm2 sterile cell culture flask. Add filtered lake water collected from each sample depth to increase the culture volume to 45 mL. Set up multiple replicates for each sampling depth if enrichments on different culture media are desired.
- Supplement culture flasks with 50X sterile solutions to a final concentration of 1X of the following culture media: Bold’s Basal Medium, BG11 and F/2. If cultivation of mixotrophic organisms is required, set-up two F/2-supplemented vessels and add 2-3 grains of sterile rice to one of the flasks.
- Incubate culture flasks at low temperature (4 °C) and low irradiance (20 μmol photons m-2 s-1) in a temperature-controlled growth incubator for at least 4 weeks in Antarctica prior to shipment to your U.S. laboratory. If isolates are desired, plate 250 μL of enrichment culture on agar plates containing appropriate growth media after 2 weeks incubation.
- For shipment to the U.S., transfer enrichment cultures to sterile 50-mL falcon tubes. Request shipment requirement of “do not freeze” and “keep cool” on shipping request. While the shipping process from Antarctica is relatively fast (2 weeks by commercial air) and highly reliable, there is a possibility of loss of fastidious organisms during the transport process. Thus, investigators may want to preserve a subsample prior to shipping so that loss of certain species during the shipping process can be assessed.
- Upon arrival in the U.S., transfer 5 mL of enrichment culture into 45 mL of appropriate growth media in a 125-mL sterile Erlenmeyer flask capped with a sterile sponge. Enrichment cultures should be maintained as standing cultures in a temperature controlled environmental growth chamber (Diurnal Growth Chamber, VWR) at 4 °C/20 μmol photons m-2 s-1. Cultures should be transferred to fresh media every 30 days.
3. Cell Lysate Extraction from Filtered Enrichments
- Filter 5 mL of enrichment culture onto a 25 mm Whatman GF/F filter using gentle vacuum (<10 psi). The filter may be stored for several weeks at -80 °C prior to assaying RubisCO activity.
- Cut the filter into 4-5 sections with sterile steel scissors and transfer the pieces with sterile forceps to a 2 mL screw cap tube filled one fifth of the way full with 0.1 mm diameter zirconia/silica beads.
- Add 1.25 mL of filter extraction buffer (50 mM bicine, 10 mM MgCl2, 1 mM EDTA, 5 mg/mL BSA, and 0.1% triton X-100; add fresh: 10 mM NaHCO3, 10 mM DTT, 3.3 mM aminocaproic acid, 0.7 mM benzamidine, pH 7.8) 13 and disrupt cells using a Minibead beater three times for 30s on speed setting 48. To prevent heating of the sample, alternate beadbeating with 1 min ice incubation.
- Centrifuge filter slurry for 2 min at 3000 x g at 4 °C to form a loose pellet of GF/F filter fibers and cell wall debris.
- Remove a 200 μL aliquot and immerse in 800 μL of ice-cold 90% acetone. Measure extractable chlorophyll (chl) a in a spectrophotometer at wavelength values of 664 nm and 647 nm 14. Transfer the supernatant to a 1.5 mL microcentrifuge tube.
- Centrifuge remaining supernatant at 4 °C for 2 min at 15,000 x g and transfer the supernatant (avoid the pellet of insoluble cellular membranes and remaining GF/F filter fibers) to a clean 1.5 mL microcentrifuge tube. This soluble cell lysate may now be used in the RubisCO carboxylase activity assay.
- Lysate may be stored at -80 °C after addition of 12.5% glycerol and flash freezing in liquid nitrogen. A loss of activity may occur after multiple freeze/thaw cycles depending on the sensitivity of the enzymes in the lysate. Therefore, activity of fresh versus frozen lysate should be tested before large scale cell lysate extractions.
4. RubisCO Carboxylase Activity Filter Assay
- Transfer 125 μL of soluble lysate to a 15 mL screw cap microcentrifuge tube (cap not needed) that has been chilled on ice. For negative control samples, add 125 μL soluble lysate to 15 mL screw cap microcentrifuge tube, but do not add substrate (in step 4.7). Run duplicate reactions for all samples and negative controls.
- Prepare 10 mL of assay buffer fresh (50 mM Bicine-NaOH, pH 8.0, 50 mM NaHCO3, 25 mM MgCl2) and chill on ice. Aliquot enough of the assay buffer for all of the samples and negative controls (100 μL per sample/control plus 100 μL extra) to a 5 mL tube. Perform all subsequent steps under a fume hood.
- Add 40 μL NaH14CO3 (500 μCi/mL) per mL of assay buffer to the aliquoted buffer and keep the buffer on ice.
- Remove 10 μL of the assay buffer containing 14C and dilute it in 1 mL of assay buffer. After inverting to mix, add 100 μL of the 1:100 diluted mixture to 3 mL of Bio-Safe II scintillation counting cocktail in a scintillation vial. Invert to mix. Scintillation counts (Beckman LS6500) should be over 18,000 cpm before continuing with the assay.
- When enough NaH14CO3 has been added to the assay buffer, move the reaction tubes to a dry bath preset to 25 °C and incubate for 3-5 mins.
- Add 100 μL assay buffer to the lysates and incubate for 5 mins at 25 °C.
- Add 20 μL of 15 mM Ribulose bisphosphate, to samples and allow the reaction to proceed for 5-10 mins at 25 °C.
- Add 100 μL propionic acid (100%) (Fisher) to stop the reaction. Centrifuge for 1 hour at 2,000 x g to exhaust unincorporated 14C.
- Transfer the total volume of the samples to scintillation vials containing 3 mL Bio-Safe II scintillation counting cocktail. Determine cpm of acid stable end products by scintillation counting (B. Witte, R. Tabita personal communication).
- Calculate RubisCO activity rates on a chl a basis 14.
5. Representative Results
We utilized enrichment culturing to isolate cold-adapted phototrophic and mixotrophic protists residing in the Antarctic Lake Bonney. To capture a greater diversity of organisms, we tested three growth media types: Bold’s Basal Medium (BBM) 15, F/2-Si Marine Medium 16, 17 and BG11 Medium 18. Visual inspection of the enrichment cultures by light microscopy revealed that the cultures were dominated by phototrophic protists (indicated by the presence of chlorophyll pigment in most cells) and harbored a variety of cell morphologies depending on the sampling depth from which the inoculum was taken and the type of media used in the culture (Fig. 1).
We measured maximum rates of carboxylase activity catalyzed by the enzyme RubisCO as a proxy for carbon fixation potential. Chl a abundance was also monitored as an estimate of phototrophic protist biomass (Fig. 2). Despite cultivation of all cultures under the same temperature/light regime (ie. 4 °C/20 μmol m-2 s-1), carbon fixation potential and phototrophic biomass varied dramatically between the enrichment cultures. Maximum RubisCO activity was observed in enrichment cultures growing in either BBM (Enrichments 4 and 6) or BG11 (Enrichments 13 and 14) growth media, while cultures enriched on F/2 growth medium (Enrichments 19, 21, 23) exhibited a 4- to 34-fold lower maximum carboxylase activities. These differences were not due to lower biomass levels in the F/2 cultures, as carboxylase activity was expressed on a Chl a basis. Moreover, RubisCO activity did not correlate with chl a concentration (r = -0.023, p>0.1; Fig. 2, inset). The BBM cultures inoculated with lake water from 6 m and 15 m (Enrichments 4 and 5) had the highest chl a levels, while all other cultures exhibited relatively low chl a, regardless of RubisCO enzyme activity (Fig. 2).
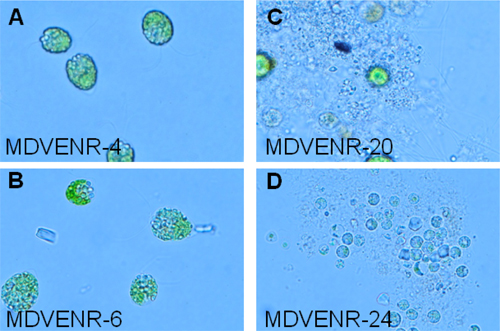
Figure 1. Representative microbial eukaryote enrichment cultures which have selected for the growth of different organisms. Identity of the culture is given in the lower left of the panels. All images represent light micrographs generated using oil immersion at 1000X magnification.
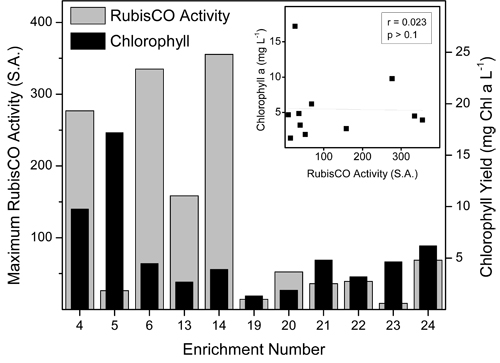
Figure 2. Potential carbon fixation capacity and extractable chlorophyll a levels in Antarctic microbial eukaryote enrichment cultures isolated from Lake Bonney and grown on various culture media. See Table 1 for culture details. Inset: Pearson correlation between chlorophyll a abundance and carbon fixation potential in Antarctic microbial eukaryote enrichment cultures.
