1. Preparation of Algal Material
- Ostreococcus cells are cultured in artificial seawater (ASW). Sea salts (typically about 40 grams per liter) are dissolved in deionised water to a salinity of 30 ppt, as measured using a salinity meter. Enrichment medium11,12, trace metal elements and vitamins are added as described in Tables 1-3. The media is then filter-sterilised through a 0.22 μm filter.
- For maintenance, cells of Ostreococcus strain OTTH95 are sub-cultured aseptically at a dilution of 1 /100 in fresh ASW every 7 days and grown under constant light in a plant growth incubator fitted with Moonlight Blue filter. The light intensity should be close to 20 μmol m-2 s-1 and temperature is maintained at 20 °C. Cells do not require constant agitation, but are shaken once every 2 to 3 days to prevent aggregation.
- For each transformation, 50 ml of cells are required at a cell density of 20-30 x 106 ml-1, which should be attained 5-7 days following sub-culturing. Approximate cell density and axeny can be determined using either flow cytometry or a haemocytometer at a minimum of x40 magnification.
2. Electroporation
- Prepare DNA for the transformation. For each transformation, 5 μg of pure, linearised plasmid DNA is required at a concentration of 1 μg/μl in sterile deionised water. To obtain this DNA, the authors recommend using a Qiagen midi prep kit, though other methods might work equally well. Digest the product with an enzyme that cuts in the backbone of the vector used, but not in the transgene or selection gene. Further purify and concentrate the resulting linear DNA by ethanol precipitation, and resuspend the product in the appropriate volume of high-quality sterile deionised water.
- Prepare microcentrifuge tubes containing 5 μg DNA for each transformation. A control with no DNA is necessary for each cell line to be transformed. Keep these tubes on ice, together with a 2 mm electroporation cuvette for each transformation.
- Prepare 2.2 ml of resuspension buffer per transformation. Dissolve Sorbitol to a 1 M solution in ddH2O, add 0.1 % pluronic acid F68 and filter-sterilise.
- Add pluronic acid F68, to a final concentration of 0.1% to the cells, and pellet them for 10 minutes at 8000x g at 10 °C in a 50 ml tube with conical bottom. Immediately resuspend the cells in 1 ml of resuspension buffer by pipetting up and down, and transfer to a microfuge tube. Spin down for 10 minutes at 8000 x g at 10 °C, and working quickly, repeat this wash step once more.
- With a cut tip, resuspend each final pellet in 40 μl of resuspension buffer. Add 40 μl of the resuspended cells to every tube of linearised DNA on ice, mix gently and transfer to the electroporation cuvette.
- Put the cuvette in the electroporation machine. Change the settings to 6 kV cm-1, 600 Ω, and 25 μF. Electroporate the cells.
- Incubate the electoporated cells in the cuvettes at room temperature for 10 minutes, and use that time to prepare the tissue culture flasks. Label them and add 30 ml of fresh ASW to each. Take 1 ml out of each flask and gently add it to the corresponding cuvette. Incubate for 2 minutes and gently remove the ASW, now containing the globule of cells and gently and slowly pipette directly into the ASW in the culture flask.
- Cells typically remain in a globule. Take care not to shake or disturb the aggregated cells at this moment. Allow the cells to recover in the incubator for 1-2 hours, then resuspend by shaking the flasks. At this time, the cells should resuspend freely and no clumps should be visible. Leave the cultures to recover overnight in the incubator.
3. Inclusion of Cells on Plates in Semi-solid Medium
- The next day, autoclave a solution of 2.1% low melting point agarose in ddH2O in a bottle containing a stirring rod. Keep molten at 65-90 °C in a larger beaker containing water on a heating magnetic stirrer. For each transformation prepare 8 Petri dishes (55 mm diameter) and 8 x 15 ml tubes each containing 9 ml of ASW plus the required selection. If using Nourseothricin or G418, use 2 mg/ml.
- Collect the transformed cells from the incubator. Working in a sterile flowhood, add 1 ml of boiling LMP agarose to the 9 ml in one of the tubes. Close the tube, and mix by inverting. Then add 0.5 ml of freshly transformed cells, quickly mix and pour into the plate. Repeat this process with the remaining 7 tubes and then proceed to the next transformation flask.
- Leave the plates open in the flow hood for an hour, for the agarose to set. Then close the plates and transfer them to large square Petri dishes, which will hold 4 plates each. Seal the square plates with parafilm. Note that the plates will not set completely, and as a result, the gel is very fragile. Care should be taken not to break the gel when handling the plates. Place all square plates in the incubator.
4. Selection of Transformed Colonies
- Colonies should appear after 10 to 21 days on the transformation plates, but not on the mock transformed plates. To pick colonies use a 200 μl pipette with cut-off tips. Simply select free colonies and suck out the green colony. Take care not to include any cells from neighbouring colonies.
- Transfer the cells to 2 ml of liquid medium containing the selection, in a 24 wells plate. When using Nourseothricin, use 1.75 mg/ml. Mix by pipetting up and down. Select 24-50 colonies per transformation. Seal the plates with parafilm and transfer to the incubator.
- After a week, transfer 100 μl of each well to 2 ml fresh ASW with selection in a 24 wells plate and grow for an additional 7 days. Surviving cell lines can be used for further studies. Stable integration into the genome and insertion number should be analysed using PCR and Southern Blot. Genomic DNA can be obtained for these purposes using any down-scaled standard method for plant DNA extraction.
5. Representative Results
Cells split 1 / 100 reached a density of 25 million cells per milliliter after growing for 7 days. Electroporation of the cells with the appropriate settings resulted in a time-constant between 10 and 14 milliseconds. When cells are transferred to medium in culture flasks, a globule of cells should form. When lightly shaken after an hour, the cells should easily resuspend. Inclusion of 1 ml transformed cells into 0.2% LMP agar should result in a consistent but only semi-solid gel. Colonies should appear after 10 to 21 days. When transforming 5 μg of linearised DNA using the protocol above, 50-100 colonies per transformation plate were typically expected, versus none on the negative control plates. ~80% of colonies picked were positively selected by antibiotic resistance in liquid medium and were used in subsequent studies.
| Final concentration | Stock solution | Volume for 1 liter | |
| NaNO3 | 8.83 x 10-4 M | 75 g/L dH2O | 1 mL |
| NH4Cl | 3.63 x 10-5 M | 2.68 g/L dH2O | 1 mL |
| β-glycerophosphate | 1 x 10-5 M | 2.16 g/L dH2O | 1 mL |
| H2SeO3 | 1 x 10-8 M | 1.29 mg/L dH2O | 1 mL |
| Tris-base(pH 7.2) | 1 x 10-3 M | 121.1 g/L dH2O | 1 mL |
| K trace metal solution | Table 2 | 1 mL | |
| f/2 vitamin solution | Table 3 | 0.5 mL |
Table 1. Medium constituents for growth of O. tauri11, 12. Make up a total volume of 1 liter of artificial seawater to a salinity of 30 ppt, and add the components above from stock solutions as indicated. Filter-sterilise the medium and use within a week. Stocks of all compounds except the vitamin solution can be pooled to add 6 ml of this solution for every liter of medium.
| Final concentration | Stock solution | Amount for 1 liter | |
| Na2EDTA • 2H2O | 1 x 10-4 M | 41.6 g | |
| FeCl3 • 6H2O | 1 x 10-5 M | 3.15 g | |
| Na2MoO4 • 2H2O | 1 x 10-8 M | 6.3 g/L dH2O | 1 mL |
| ZnSO4 • 7H2O | 1 x 10-9 M | 22.0 g/L dH2O | 1 mL |
| CoCl2• 6H2O | 1 x 10-9 M | 10.0 g/L dH2O | 1 mL |
| MnCl2 • 4H2O | 1 x 10-8 M | 180.0 g/L dH2O | 1 mL |
| CuSO4• 5H2O | 1 x 10-8 M | 9.8 g/L dH2O | 1 mL |
Table 2. Trace metal solution11,12. Make up to a total of 1 liter, in dH2O, and heat to dissolve. Aliquot the solution and freeze for storage. Final concentrations indicated refer to the final medium, not the trace metal solution.
| Final concentration | Stock solution | Amount for 1 liter | |
| Vitamin B12 | 1 x 10-10 M | 1 g/L dH2O | 1 mL |
| Biotin | 1 x 10-9 M | 0.1 g/L dH2O | 10 mL |
| Thiamine • HCl | 1 x 10-7 M | 200 mg |
Table 3. f/2 vitamin solution11. Make up to a total of 1 liter in dH2O, filter-sterilise, aliquot and freeze. Final concentrations indicated refer to the final medium, not the vitamin solution.
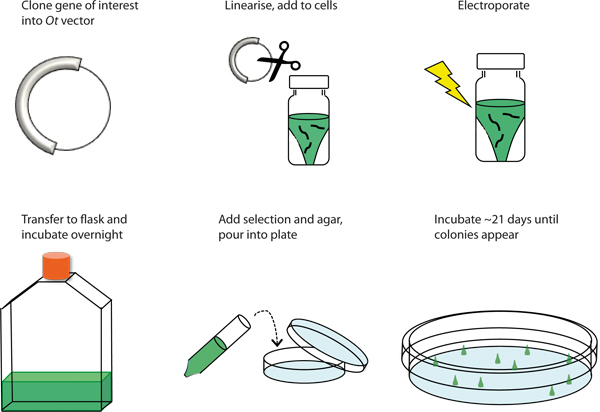
Figure 1. Graphical overview of the transformation procedure. Schematic representation of the procedure to genetically transform Ostreococcus tauri by electroporation.
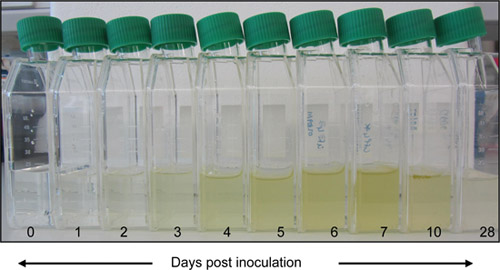
Figure 2. Culture growth. Cells are sub-cultured aseptically at a dilution of 1 /100 in fresh ASW every 7 days and grown under constant light in a plant growth incubator fitted with Moonlight Blue filter. The light intensity should be close to 20 μmol m-2s-1 and temperature is maintained at 20 °C Cells do not require constant agitation, but are shaken once every 2 to 3 days to prevent aggregation.
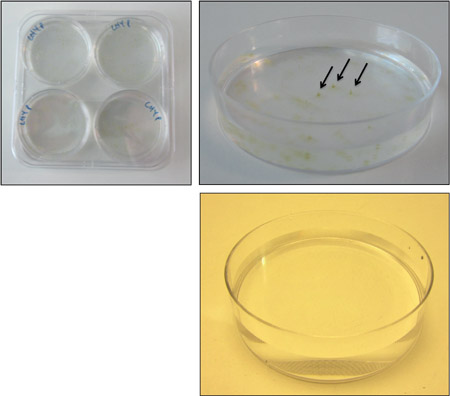
Figure 3. Colony formation on 0.2% LMP agarose. Transfer 4 plates to a large square Petri dish, and seal with parafilm (top left). When colonies have formed, simply select free (ideally conically shaped) colonies (top right) and suck them out of the plate with a p200 pipette. There should be no colonies on the negative control (bottom right).
